Há um número significativo de compostos endócrinos (EDC) que estão entre as substâncias mais perigosas em nosso ambiente. Estes são principalmente compostos estrogênicos que contaminam a água dos recursos naturais. A diversidade química das substâncias pertencentes ao grupo dificulta os testes para sua presença, uma vez que diferentes métodos analíticos são necessários para sua detecção. Com base em sua estrutura química é muito difícil determinar se uma substância é realmente capaz de agir como um estrogênio. Além disso, essas substâncias nunca estão presentes de forma pura no ambiente, de modo que seus efeitos podem ser afetados por outros compostos, também1. Esse problema pode ser resolvido por métodos de detecção de efeitos, como o uso de organismos biomonitor/bioindicadores que apresentam efeitos estrogênicos2,,3,,4,,5.
Recentemente, uma variedade de sistemas de teste baseados em células6 eleveduras 2,3 foram desenvolvidos para detectar efeitos estrogênicos. No entanto, estes são geralmente apenas capazes de detectar a ligação da substância ao receptor de estrogênio2,3. Além disso, são incapazes de modelar processos fisiológicos complexos no organismo, ou detectar fases sensíveis ao hormônio das fases da vida; assim, muitas vezes levam a resultados falsos.
Sabe-se que certos genes reagem sensivelmente ao estrogênio em organismos vivos7. A detecção de produtos genéticos por métodos de biologia molecular também é possível no nível de proteína ou mRNA8,,9, mas geralmente envolve sacrifício animal. As leis de proteção animal tornaram-se mais rigorosas, e há uma demanda crescente por sistemas de teste alternativos que minimizem o número e o sofrimento dos animais usados em experimentos ou a substituição do modelo animal por outro modelo de sistema10. Com a descoberta de proteínas fluorescentes e a criação de linhas biomarcadoras, as tecnologias transgênicas fornecem uma boa alternativa11. Com essas linhas, a ativação de um gene sensível ao estrogênio pode ser testada in vivo.
Entre os vertebrados, destaca-se o potencial dos peixes na avaliação de risco ambiental. Eles oferecem muitas vantagens sobre os modelos de mamíferos: sendo organismos aquáticos, eles são capazes de absorver poluentes através de todo o seu corpo, produzir um grande número de descendentes, e algumas de suas espécies são caracterizadas pelo curto tempo de geração. Seu sistema endócrino e processos fisiológicos mostram grandes semelhanças com outros vertebrados e até mesmo com mamíferos, incluindo humanos12.
Vários genes para a detecção de efeitos estrogênicos em peixes também são conhecidos. Os mais importantes são os receptores de estrogênio aromatase-b, choriogenina-H e vitellogenina (vtg)7,13. Recentemente, várias linhas biosensoras produtoras de estrogênio também foram criadas a partir de modelos de peixes utilizados em laboratório, como a de zebrafish (Danio rerio)4,5,14,15,16,17. A principal vantagem do zebrafish na criação de linhas biosensoras é o corpo transparente dos embriões e larvas, pois o sinal fluorescente do repórter pode então ser facilmente estudado in vivo sem sacrificar o animal10. Além da proteção animal, também é uma característica valiosa, pois permite estudar a reação do mesmo indivíduo em diferentes momentos do tratamento18.
Esses experimentos usam um repórter de vitellogenina linhazebrafish transgênica 15. A construção transgênica utilizada para o desenvolvimento de Tg(vtg1:mCherry) tem um longo (3,4 kbp) promotor de vitellogenina natural-1. O receptor de estrogênio (ER) é uma proteína melhorador ativada por ligantes que é um representante da superfamília esteroide/receptor nuclear. O ER se liga a sequências específicas de DNA chamadas elementos de resposta de estrogênio (EREs) com alta afinidade e transativa a expressão genética em resposta ao estradiol e outras substâncias estrogênicas, de modo que quanto mais ERE no promotor causa uma resposta mais forte19. Existem 17 locais de ERE na região promotora da construção transgênica Tg(vtg1:mCherry) e espera-se que eles imitem a expressão do gene vtg nativo15. Há uma expressão contínua do sinal fluorescente em fêmeas sexualmente amadurecidas. No entanto, no sexo masculino e embrião a expressão no fígado só é visível após o tratamento com substâncias estrogênicas(Figura 1).
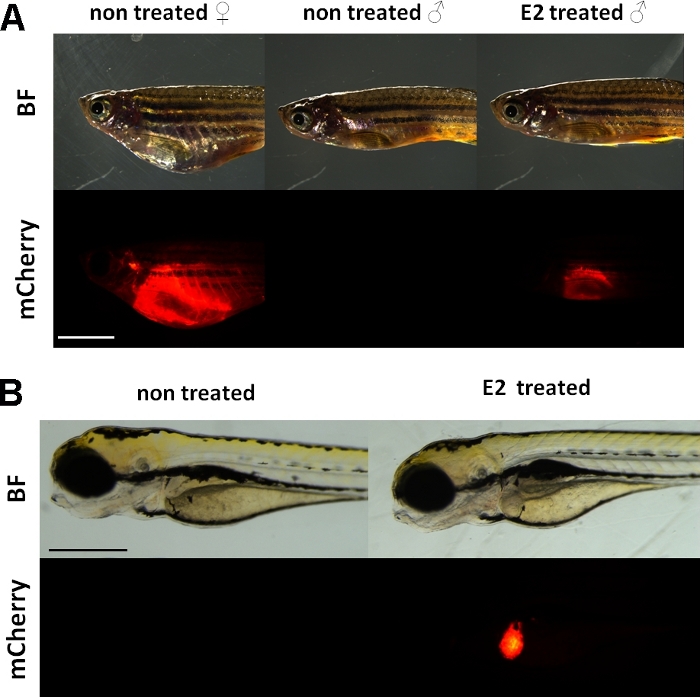
Figura 1: Sinal fluorescente vermelho no fígado de vtg1:mCrestra-se peixe-zebra adulto transgênico e 5 embriões dpf, após indução de 17-ß-estradiol (E2). No feminino e no sexo masculino tratado com E2 (25 μg/L tempo de exposição:48hrs) a fluorescência forte do fígado é visível mesmo através da pele pigmentada. Nenhum sinal fluorescente é visível em macho não tratado(A). Após a indução do E2 (50 μg/L tempo de exposição: 0-120 hpf), também pode ser observado um sinal fluorescente vermelho no fígado de 5 embriões dpf, que não é visível em embriões de controle(B). Embora o sinal fluorescente esteja continuamente presente em fêmeas adultas, principalmente machos e embriões da linha são adequados para detectar efeitos estrogênicos. (BF: campo brilhante, mCherry: visualização do filtro fluorescente vermelho, imagens simples simples, barra de escala A: 5mm, barra de escala B: 250 μm) Clique aqui para ver uma versão maior desta figura.
Semelhante à vitellogenina endógena, o repórter mCherry só é expresso no fígado. Como a vitellogenina só é produzida na presença de estrogênio, não há sinal fluorescente nos controles. Como a expressão está apenas no fígado, a avaliação dos resultados é muito mais fácil15.
A sensibilidade e a usabilidade dos embriões desta linha têm sido investigadas em várias misturas de compostos estrogênicos e também em amostras ambientais15,,20, e na maioria dos casos foram documentadas relações dose-resposta(Figura 2). No entanto, no caso de substâncias altamente tóxicas, principalmente hepatotóxias (por exemplo, zearalenone), apenas um sinal fluorescente muito fraco pode ser visível no fígado de embriões tratados e o sinal fluorescente de intensidade máxima causado pode ser alcançado dentro de uma faixa de concentração muito pequena, o que dificulta a criação de relações dose-efeito20.
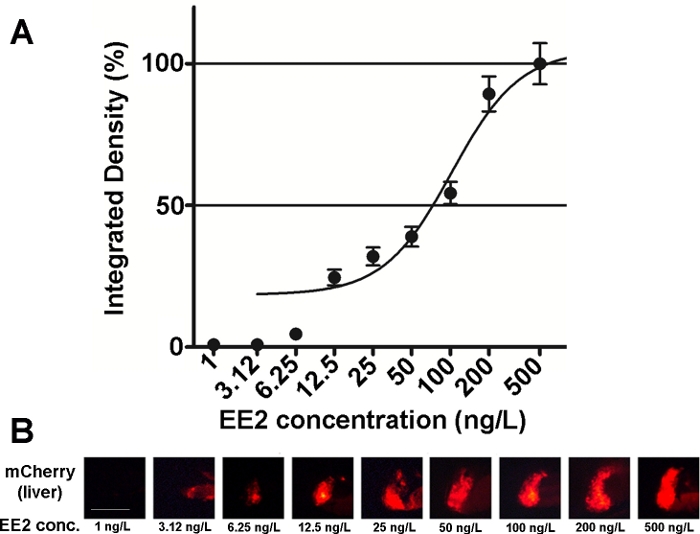
Figura 2: Diagrama de dose-resposta (A) e imagens fluorescentes (mCherry) do fígado (B) expostas a 17-α-ethynilestradiol (EE2), em 5 dpf vtg1:mAsinas decherry. Os resultados são expressos como densidade integrada gerada a partir da força do sinal e do tamanho da área afetada (±SEM, n = 60). 100% refere-se ao máximo observado. A intensidade do sinal fluorescente aumentou gradualmente com a concentração. Barra de escala = 250 μm. Clique aqui para ver uma versão maior desta figura.
Existem várias substâncias estrogênicas presentes no ambiente, tais como 17-β-estradiol (concentração ambiental: 0.1-5.1 ng/L)21, 17-α-ethynylestradiol (concentração ambiental: 0.16-0.2 μg/L)22, zearalenone (concentração ambiental: 0,095-0,22 μg/L)23, bisfenol-A (concentração ambiental: 0,45-17,2 mg/L)24. Ao testar essas substâncias de forma ativa pura com a ajuda de embriões transgênicos mCherry, as concentrações de efeitos mais baixas observadas (LOEC) para detecção de sinais fluorescentes foram de 100 ng/L para 17-ß-estradiol, 1 ng/L para 17-α-ethynilestradiol, 100 ng/L para zearalenone e 1 mg/L para bisfenol-A (tratamento de 96-120 hpf), que está muito próximo ou dentro da faixa de concentrações ambientais das substâncias15. A linha transgênica Tg(vtg1:mCherry) pode ajudar a detectar estrogenicidade em amostras de águas residuais após exposição direta. A linha é tão sensível quanto o teste de estrogênio de levedura comumente usado, o ensaio de estrogênio bioluminiscente (BLYES)15. Com a ajuda desta linha, os efeitos protetores dos beta-ciclodextrinas contra a toxicidade induzida por zearalenona foram confirmados usando misturas químicas20.
Em um relatório recente, o uso in vivo da linha transgênica foi demonstrado com a ajuda de dois metabólitos zearalenone estrogênico (ZEA), α- e β-zearalenol (α-ZOL e β-ZOL)25. A linha de base do protocolo é apropriada para estudar os efeitos estrogênicos de vários compostos ou amostras ambientais em embriões Tg(vtg1:mCherry).
