Drevet af den stigende anvendelse af næste generations sekventeringsteknikker kan rumlig opløsning i udviklingsbiologi øges. Moderne undersøgelser har til formål at dissekere væv ned til specialiserede celletyper, hvis ikke enkeltcelleniveau1,2,3,4. Med henblik herpå er der i løbet af de sidste 50 år udviklet en overflod af forskellige metoder (se figur 1A)5,6,7,8,9,10,11,12,13,14,15.
Mange værktøjer i plantevidenskab har været tilpasninger af teknikker, der var banebrydende inden for dyreforsøg. Det er ikke tilfældet for den metode, vi indfører i detaljer her. I 2005, udstyret med en stærk baggrund i protein oversættelse, Bailey-Serres Lab satte sig for at ingeniør ribosomale proteiner til efterfølgende affinitet rensning16. Således kunne de undgå tidskrævende og arbejdskrævende polysome profilering, som er baseret på ultracentrifugering med en saccharose gradient og blev brugt til at vurdere oversætte ribosomer siden 1960’erne17,18. Metoden er siden blevet omtalt som translationel ribosom affinitetrensning (TRAP)16. Efter vellykkede oversættelsesundersøgelser i planter, Heiman et al. tilpasset TRAP for dyr19 og andre udvidet sin ansøgning til gær20, Drosophila21, Xenopus22 og zebrafisk23,24.
Selv om genetisk modifikation af modelsystemet er en forudsætning for TRAP, som begrænser dets anvendelse til arter, der kan underkastes genetisk transformation, kan man samtidig udnytte denne indvending til at målrette delmængder af celler, der er af særlig interesse og ellers yderst vanskeligt at isolere fra det intakte væv/organ25 (f.eks. stærkt forgrenede dendritiske celler i en musehjerne eller svampehyphae i inficeret plantevæv). I planter holdes alle celler på plads via cellevægge, der danner grundlag for det hydrostatiske skelet26. For at frigøre en plantecelle fra denne matrix har forskerne enten fysisk skåret cellen ud af det omgivende væv gennem lasercapture microdissection (LCM)27 eller udført enzymatisk fordøjelse af cellevæggene28. Blandt de sidstnævnte celler, såkaldte protoplasts, er populationen af interesse fluorescerende mærket og kan adskilles via fluorescens-aktiveret cellesortering (FACS)7. LCM kræver normalt en prøve, der skal fastsættes og indlejres i voks, hvilket i sidste ende forringer kvaliteten af sin RNA29. FACS-baserede metoder giver høj kvalitet RNA, men processen med protoplasting selv introducerer forskelle i genekspression30 og væv med modificerede og tykke sekundære cellevægge er notorisk vanskelige at behandle. Desuden antages mange udviklingsprocesser i planter at være afhængige af mekanisk transmitterede signaler, og derfor er cellevæggens integritet af afgørende betydning31. To metoder, der bruger en genvej til at omgå celleisolation ved at operere på niveauet af kerner, er fluorescensaktiveret nuklear sortering (FANS) og isolering af kerner mærket i bestemte celletyper (INTAKT). Som i TRAP, de bruger celletype-specifikke initiativtagere til at markere kerner, der efterfølgende bliver beriget via sortering eller trække ned, henholdsvis8,15. En stor udfordring for alle disse tilgange er at få tilstrækkeligt RNA-materiale fra delmængder af celler i et væv. Da TRAP kun fanger en brøkdel af de cellulære RNA’er, er prøveindsamling en betydelig flaskehals. Derfor er der behov for særligt følsomme biblioteksforberedelsesprotokoller for at producere data af høj kvalitet fra lave inputmængder.
Siden oprettelsen er TRAP enten blevet anvendt i kombination med DNA-mikroarrays eller, da sekventeringsomkostningerne er faldet betydeligt i de senere år, RNA-seq10,32,33. En lang række forskningsspørgsmål er allerede blevet belyst som gennemgået i Sablok et al.34. Vi er overbeviste om, at flere rapporter vil følge i de kommende år, da teknikken er meget alsidig, når man kombinerer forskellige initiativtagere til at målrette specifikke celletyper. Til sidst, Dette vil ske selv i en ukuelig måde, og kan kombineres med sondering plantens reaktion på mange biotiske og abiotiske stressfaktorer. Derudover, hvor stabile transgene linjer ikke er tilgængelige, behårede rod udtryksystemer er også blevet anvendt med succes til at udføre TRAP i tomat og medicago35,36.
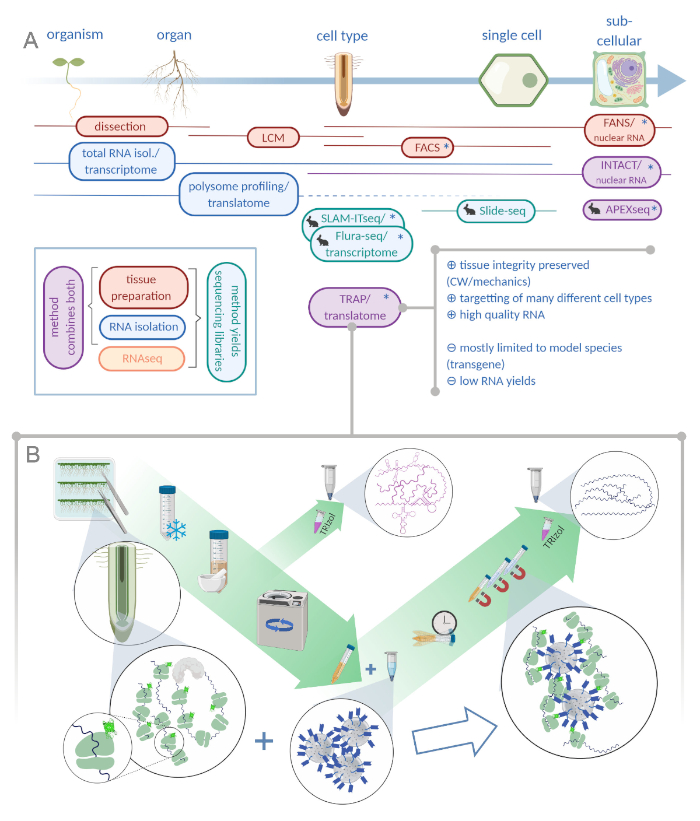
Figur 1: Oversættelse af ribosom affinitetsrensning (TRAP) supplerer analyseporteføljen “omics”. A. Stigende niveauer af analytisk præcision, ned til encellede eller endda subcellulære opløsning kan opnås ved et væld af metoder eller kombinationer heraf. Ordningen giver et overblik over de værktøjer, der i øjeblikket er til rådighed på plante- og dyreområdet. Vævssamling ved cellulær opløsning kan opnås ved protokoller som LCM eller FACS, som derefter kobles til standard transkription eller polysome profileing/translatome analyse. TRAP og INTAKT integrerer både vævsindfangning og RNA-isolation, da de er baseret på epitop-tagging. Intakt e-prøver imidlertid kun cellekerner og udgør derfor et særligt tilfælde af transskriptionsanalyse. En lille kanin ikon markerer nyudviklede metoder på dyreområdet: Mens SLAM-ITseq og Flura-seq stole på metaboliske målretning af spirende RNAs med modificerede uracil baser i celler, der udtrykker eftergivende enzym, Slide-seq gør brug af en belagt glas dias med DNA stregkoder, der giver positionelle oplysninger i cellulære område. Der anvendes en nærhedsmærkningsmetode i APEX-seq for at udtage prøver af RNA’er i specifikke subcellulære rum. Navnlig kræver øget opløsning ofte generering af transgent materiale (stjerner), og disse metoder anvendes således overvejende til modelarter. TRAP er specielt velegnet til plantevidenskabelige undersøgelser, der involverer cellevæg (CW) eller mekaniker signalering samt cellearter, der er vanskelige at frigive fra deres CW matrix. B. Detaljerede vådlaboratorietrin i TRAP-proceduren: Kimplanter, der udtrykker GFP-mærket ribosomprotein i forskellige celletyper (f.eks. rodendodermis), dyrkes på petriskåle i syv dage og rodmateriale høstet ved snapfrysning. En total RNA-kontrolprøve indsamles fra det homogeniserede rå ekstrakt, før det pelleteres i snavset via centrifugering. Magnetiske anti-GFP perler tilsættes til det klare ekstrakt for at udføre immunudfældning. Efter inkubation og tre vasketrin opnås det polysomeassocierede RNA (TRAP/polysome RNA) direkte via fenol-chloroform ekstraktion. LCM: mikrodissektion af laserfanger, FACS/FANS: fluorescensaktiveret celle/nuklear sortering APEX-seq: metode baseret på manipuleret ascorbatperoxidase, INTAKT: isolering af kerner mærket i specifikke celletyper, SLAM-ITseq: thiol(SH)-linked alkylering for metabolisk sekventering af RNA i væv, Flura-seq: fluorouracil-mærket RNA sekventering (Oprettet med Biorender.com) Klik her for at se en større version af denne figur.
Målet med denne artikel er at give en detaljeret beskrivelse af TRAP-metoden, at fremhæve kritiske trin og at give vejledning til en mulig metode til forberedelse af biblioteket.
Et generisk TRAP-eksperiment vil hovedsagelig bestå af følgende trin (se også figur 1B):(1) Fremstilling af plantemateriale, herunder kloning af ribosomt-taggingkonstruktion, transgene linjeproduktion og -udvælgelse, dyrkning og opsamling af frø, sterilisering og plettering samt stressanvendelse/-behandling (valgfri) og vævshøst; 2) immunrensning, herunder vævshomogenisering og rydning af råekstrakt, perlevask og immunrensning samt vasketrin 3) RNA-ekstraktion og kvalitetsvurdering og (4) biblioteksforberedelse.
Arabidopsis-roden har været et modelsystem til undersøgelse af planteudvikling lige siden dens indførelse som modelanlæg37,38. Her er anvendelsen af TRAP fremvist i forbindelse med anlæggetlateral rod udvikling. I planter, opbygningen af hele rodsystemet er afhængig af udførelsen af dette program og er derfor meget vigtigt for overlevelsen af organismen39. I Arabidopsisstammer laterale rødder fra pericyclevæv , der ligger ved siden af xylem-karog derfor kaldes xylempolpericycle (XPP; se figur 2C)40. Nogle XPP-celler, som er placeret dybt inde i roden, erhverve en grundlægger celleidentitet og, på en lokal hormonelle udløse, begynder at formere sig ved hævelse og dividere anticlinally41. Men på grund af tilstedeværelsen af en stiv cellevæg matrix, denne proces udøver mekanisk stress på det omgivende væv. Især er den overliggende endodermis påvirket, da det er i vejen for den laterale rodvækst akse42,43,44. Faktisk vil den nydannende primordium nødt til at vokse gennem den overliggende endodermis celle (Figur 2C2), mens cortex og epidermis celler er bare skubbet til side for primordium til endelig at dukke45,46. Nylige arbejde i vores laboratorium har vist, at endodermis aktivt bidrager til at imødekomme spredning i perihjulet. Målrettet blokering af endodermal hormonel signalering er tilstrækkelig til at hæmme selv den allerførste division i XPP-cellerne47. Derfor pericycle-endodermis kommunikation udgør en meget tidlig checkpoint for lateral rod udvikling i Arabidopsis. Det vides dog ikke, hvordan denne krydstale udføres. For at opklare dette mysterium valgte vi TRAP-seq-tilgangen til at målrette XPP og endodermale celler. At berige for celler i den laterale rod program, vi efterlignede hormonelle udløse ved eksogent anvende en auxin analog (1-naphthaleneeddikesyre, NAA)48, som samtidig lov til at tidsmæssigt løse den indledende fase af lateral roddannelse.
