Spinto dalla crescente applicazione delle tecniche di sequenziamento di prossima generazione, la risoluzione spaziale nella biologia dello sviluppo potrebbe essere aumentata. Studi contemporanei mirano a sezionare i tessuti verso il basso per tipi di cellule specializzate, se non il livello a cella singola1,2,3,4.4 A tal fine, negli ultimi cinquant’anni è stata ideata una pletora di metodi diversi (vedi Figura 1A)5,6,7,8,9,10,11,12,13,14,15.
Molti strumenti nella scienza delle piante sono stati adattamenti di tecniche che sono state introdotte nella ricerca animale. Questo non è il caso per il metodo che stiamo introducendo in dettaglio qui. Nel 2005, dotato di un forte background nella traduzione delle proteine, il Bailey-Serres Lab ha deciso di progettare proteine ribosomiche per la successiva purificazione dell’affinità16. Così, potrebbero evitare la profilazione polisografica lunga e laboriosa, che si basa sull’ultracentrifugazione con un gradiente di saccarosio ed è stato utilizzato per valutare i ribosomi traslitteranti dal 196017,18. Da allora il metodo è stato indicato come trap (Affinity purificazione)16. Dopo studi translatomi di successo in piante, Heiman et al. adattato TRAP per gli animali19 e altri esteso la sua applicazione al lievito20, Drosophila21, Xenopus22 e pesce zebra23,24.
Anche se la modificazione genetica del sistema modello è un prerequisito per il TRAP, che limita la sua applicazione alle specie suscettibili di trasformazione genetica, si può allo stesso tempo sfruttare questa obiezione per indirizzare sottoinsiemi di cellule che sono di particolare interesse e altrimenti estremamente difficili da isolare dal tessuto intatto / organo25 (ad esempio, cellule dendritiche altamente ramificate in un cervello di topo o iphae fungini intessuto vegetale infetto). Nelle piante, tutte le cellule sono tenute in posizione attraverso pareti cellulari che costituiscono la base dello scheletro idrostatico26. Per liberare una cellula vegetale da questa matrice, gli scienziati hanno fisicamente tagliato la cellula dal tessuto circostante attraverso la microdissezione di cattura laser (LCM)27 o eseguito la digestione enzimatica delle pareti cellulari28. Tra queste ultime cellule, le cosiddette protoplasts, la popolazione di interesse è fluorescente etichettata e può essere separata tramite lo smistamento cellulare attivato dalla fluorescenza (FACS)7. LCM di solito richiede un campione da fissare e incorporare nella cera, che alla fine deteriora la qualità del suo RNA29. I metodi basati su FACS producono RNA di alta qualità, ma il processo di protoplasting stesso introduce differenze nell’espressione genica30 e i tessuti con pareti cellulari secondarie modificate e spesse sono notoriamente difficili da trattare. Inoltre, si presume che molti processi di sviluppo negli impianti si basino su segnali trasmessi meccanicamente e quindi l’integrità della parete cellulare è di fondamentale importanza31. Due metodi, che utilizzano una scorciatoia per aggirare l’isolamento cellulare operando a livello di nucleii, sono lo smistamento nucleare attivato dalla fluorescenza (FANS) e l’isolamento dei nuclei taggati in specifici tipi di cellule (INTACT). Come nel TRAP, usano promotori specifici del tipo di cellula per contrassegnare i nuclei, che successivamente si arricchiscono tramite ordinamento o discesa, rispettivamente8,15. Una grande sfida per tutti questi approcci è ottenere sufficiente materiale di RNA da sottoinsiemi di cellule in un tessuto. Poiché TRAP cattura solo una frazione degli RNA cellulari, la raccolta dei campioni è un notevole collo di bottiglia. Pertanto, sono necessari protocolli di preparazione della libreria particolarmente sensibili per produrre dati di alta qualità da quantità di input basse.
Dalla sua istituzione, TRAP è stato utilizzato in combinazione con microarray di DNA o, come i costi di sequenziamento è sceso in modo significativo negli ultimi anni, RNA-seq10,32,33. Una moltitudine di domande di ricerca è già stato chiarito come esaminato in Sablok et al.34. Siamo convinti che nei prossimi anni seguiranno ulteriori rapporti, poiché la tecnica è molto versatile quando si combinano diversi promotori per indirizzare tipi di cellule specifici. Alla fine, questo sarà fatto anche in modo inducibile, e può essere combinato con sondare la reazione della pianta a molti fattori di stress biotici e abiotici. Inoltre, laddove non sono disponibili linee transgeniche stabili, sono stati utilizzati con successo anche sistemi di espressione delle radici pelose per eseguire trap in pomodoro e medicago35,36.
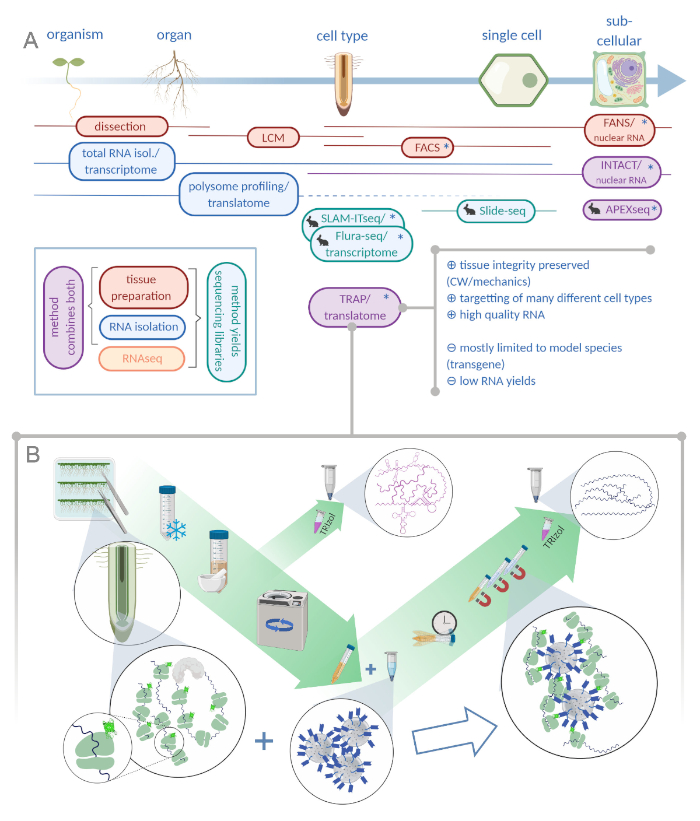
Figura 1: La traduzione della purificazione dell’affinità del ribosomiare (TRAP) integra il portafoglio di analisi “omiche”. R. L’aumento dei livelli di precisione analitica, fino alla risoluzione a cella singola o addirittura subcellulare, può essere raggiunto con una pletora di metodi o combinazioni. Il sistema fornisce una panoramica degli strumenti attualmente disponibili nel campo delle piante e degli animali. La raccolta dei tessuti a risoluzione cellulare può essere ottenuta da protocolli come LCM o FACS, che vengono poi accoppiati a trascrittoma standard o all’analisi di profilazione/traslatoma polisomico. TRAP e INTACT integrano sia la cattura dei tessuti che l’isolamento dell’RNA in quanto si basano sull’epitope-tagging. Tuttavia, INTACT campiona solo i nuclei cellulari e costituisce, quindi, un caso speciale di analisi del trascrittoma. Una piccola icona coniglio segna metodi appena sviluppati nel campo animale: Mentre SLAM-ITseq e Flura-seq si basano sul targeting metabolico di RNA nascenti con basi uracili modificate in cellule che esprimono l’enzima permissivo, Slide-seq fa uso di un vetrino di vetro rivestito con codici a barre del DNA che forniscono informazioni posizionali nella gamma cellulare. In APEX-seq è seguito un approccio di etichettatura di prossimità per campionare gli RNA in specifici compartimenti subcellulari. In particolare, una maggiore risoluzione spesso richiede la generazione di materiale transgenico (asterischi) e questi metodi sono quindi utilizzati prevalentemente per le specie modello. Il TRAP è particolarmente adatto per studi di scienze vegetali che coinvolgono la parete cellulare (CW) o la segnalazione meccanica, nonché le specie cellulari che sono difficili da rilasciare dalla loro matrice CW. B. Le fasi dettagliate del laboratorio umido della procedura TRAP: le piantine che esprimono proteine ribosomiche marcate GFP in tipi di cellule distinte (ad esempio l’endodermide delle radici) vengono coltivate sui piatti Petri per sette giorni e il materiale delle radici raccolto dal congelamento a scatto. Un campione totale di controllo dell’RNA viene raccolto dall’estratto grezzo omogeneo prima di pellere i detriti attraverso la centrifugazione. Perline magnetiche anti-GFP vengono aggiunte all’estratto eliminato per eseguire l’immunoprecipitazioni. Dopo l’incubazione e tre fasi di lavaggio, l’RNA associato al poliso (TRAP/RNA polisopo) viene ottenuto direttamente tramite l’estrazione del fenolo-cloroformio. LCM: microdissezione di cattura laser, FACS/FANS: smistamento cellulare/nucleare attivato dalla fluorescenza, APEX-seq: metodo basato su ingegnerizzato ascorbate perossidasi, INTACT: isolamento dei nuclei contrassegnati in specifici tipi di cellule, SLAM-ITseq: thiol(SH)-linked alchilirazione per il sequenziamento metabolico dell’RNA nel tessuto, Flura-seq: fluorouracil-labeled RNA sequencing (Creato con Biorender.com) Fare clic qui per visualizzare una versione più ampia di questa figura.
L’obiettivo di questo articolo è fornire una descrizione dettagliata del metodo TRAP, evidenziare i passaggi critici e fornire indicazioni per un possibile metodo di preparazione della libreria.
Un esperimento trap generico consisterà essenzialmente nei seguenti passi (vedi anche Figura 1B): (1) Preparazione del materiale vegetale, compresa la clonazione del costrutto di ribosomi, la produzione e la selezione di linee transgeniche, la coltivazione e l’accumulo di semi, la sterilizzazione e la placcatura e l’applicazione/trattamento dello stress (opzionale) e la raccolta dei tessuti; (2) immunopurificazione, compresa l’omogenesi e la bonifica dei tessuti, il lavaggio delle perline e l’immunopurificazione e le fasi di lavaggio; (3) Estrazione dell’RNA e valutazione della qualità; e (4) la preparazione della biblioteca.
La radice dell’Arabidopsis è stata un sistema modello per studiare lo sviluppo delle piante sin dalla sua introduzione come impianto modello37,38. Qui, l’applicazione di TRAP è presentata nel contesto dello sviluppo delle radici laterali dell’impianto. Nelle piante, l’accumulo dell’intero sistema radicale si basa sull’esecuzione di questo programma ed è quindi molto importante per la sopravvivenza dell’organismo39. In Arabidopsis, le radici laterali provengono dal tessuto periciclo che risiede accanto ai vasi xylem e quindi è chiamato xylem polo periciclo (XPP; vedi Figura 2C)40. Alcune cellule XPP, che si trovano in profondità all’interno della radice, acquisiscono un’identità cellulare fondatore e, su un trigger ormonale locale, iniziano a proliferare gonfiore e dividendo anticlinally41. Tuttavia, a causa della presenza di una matrice rigida della parete cellulare, questo processo esercita uno stress meccanico sui tessuti circostanti. In particolare, l’endodermide sovrastante è interessata, in quanto è nel modo dell’asse di crescita della radice laterale42,43,44. Infatti, il primordio appena formato dovrà crescere attraverso la cellula endodermide sovrastante (Figura 2C2) mentre le cellule corteccia ed epidermide sono solo spinte da parte per il primordio per emergere finalmente45,46. Recenti lavori nel nostro laboratorio hanno dimostrato che l’endodermide sta contribuendo attivamente ad accogliere la proliferazione nel periciclo. Il blocco mirato della segnalazione ormonale endodermica è sufficiente per inibire anche la prima divisione nelle cellule XPP47. Pertanto, la comunicazione pericycle-endodermis costituisce un checkpoint molto precoce per lo sviluppo della radice laterale nell’Arabidopsis. Tuttavia, non è noto come questo crosstalk viene eseguito. Per svelare questo mistero, abbiamo scelto l’approccio TRAP-seq per colpire XPP e cellule endodermiche. Per arricchire per le cellule nel programma della radice laterale, abbiamo imitato il trigger ormonale applicando esogenamente un analogo auxina (1-naftaleneacetic acid, NAA)48, che allo stesso tempo ha permesso di risolvere temporaneamente la fase iniziale di formazione della radice laterale.
