تتعرض الخلايا البشرية باستمرار لمجموعة متنوعة من العوامل الضارة الحمض النووي من أصول مختلفة. المصادر الخارجية تتكون في الغالب من التعرض للإشعاعات والمواد الكيميائية (بما في ذلك العوامل العلاجية الكيميائية وبعض المضادات الحيوية) والفيروسات، في حين أن المصادر الذاتية الرئيسية تشمل أخطاء في تكرار الحمض النووي والإجهاد التأ المؤدّس. ويمكن أن تتراوح الآثار المباشرة للتعرض للسمية الجينية من قاعدة معدلة إلى كسر مزدوج حبل الحمض النووي القاتل المحتمل ، اعتمادا على الإجهاد وجرعة التعرض. في نهاية المطاف، يمكن أن يؤدي تلف الحمض النووي غير المصلح أو غير إصلاحه إلى تراكم الطفرات وإعادة ترتيب الجينوم وعدم استقرار الجينوم ويؤدي في نهاية المطاف إلى الإصابة بالسرطان1. وقد تطورت خلايا الثدييات مسارات معقدة للتعرف على أنواع محددة من الضرر الحمض النووي2,3 وإصلاحها في الوقت المناسب, تزامن مع تطور دورة الخلية.
الإشعاع المؤين (IR) يضر الحلزون المزدوج الحمض النووي ويخلق فواصل حبلا مزدوج (DSBs)، واحدة من أكثر أشكال الضرر الضارة الحمض النووي. وMN (MRE11، RAD50، NBS1) وظائف معقدة كمستشعر من ينتهي الحمض النووي وينشط كيناز البروتين ترنح تيلانغيكتسيا تحور (ATM)4،5. بعد التنشيط الأولي من أجهزة الصراف الآلي من قبل ينتهي الحمض النووي، أجهزة الصراف الآلي يؤدي سلسلة من الأحداث DDR في موقع الاستراحة، والشروع مع حدث رئيسي، وفسفورل من البديل هيستون H2AX6. H2AX الفوسفور على بقايا S139 ينشط في γH2AX ، تمتد المناطق حتى megabases حول آفة الحمض النووي6،7،8،9. هذا الحدث يزيد من إمكانية الحصول على الحمض النووي، مما يؤدي إلى تجنيد وتراكم البروتينات إصلاح الحمض النووي الأخرى7. لأن γH2AX هو بزفير وعلى وجه التحديد المستحثة DSBs المحيطة، فإنه يمكن تصور بسهولة باستخدام أجسام مضادة محددة، ويستخدم عادة كعلامة بديلة لDSBs في مجال إصلاح الحمض النووي. بمجرد الإشارة إلى الكسر ، تقوم الخلايا بتنشيط مسارات إصلاح الحمض النووي ومعالجة تلف الحمض النووي. البروتين MDC1 (وسيط الحمض النووي الضرر نقطة تفتيش البروتين 1) يربط مباشرة γH2AX10, يتفاعل مع أجهزة الصراف الآلي11 وأيضا مع NBS112,13. وهو يساهم في زيادة تركيز مركب MRN في DSB وبدء حلقة تغذية مرتدة إيجابية الصراف الآلي. تتم إزالة γH2AX بسرعة بمجرد إصلاح الكسر ، وبالتالي ، مما يسمح بمراقبة إزالة DSB. تليها المجهر، وانخفاض في γH2AX مع مرور الوقت يوفر قياس غير مباشر من فواصل المتبقية وكفاءة إصلاح الحمض النووي.
يمكن للخلايا النوى إصلاح DSBs من قبل عدة مسارات، وهما الرئيسية التي يجري غير متجانسة نهاية الانضمام (NHEJ) وإعادة التركيب المتماثل (الموارد البشرية) (الشكل 1). NHEJ أساسا ربط الحمض النووي مزدوجة حبلا ينتهي دون استخدام homology الموسعة وتعمل في جميع أنحاء دورة الخلية14،15. الموارد البشرية تصبح الغالبة خلال مراحل S و G2 ، وقمعها على خلاف ذلك ، لأنه يتطلب chromatid الشقيقة كقالب متجانس لإصلاح14،16. اختيار المسار بين NHEJ و الموارد البشرية لا يعتمد فقط على القرب المادي من الكروماتيد الشقيقة، ولكن أيضا على تمديد استئصال نهاية الحمض النووي17، مما يمنع NHEJ.
إصلاح DSB المعتمد على Homology يبدأ عن طريق التحلل النووي من حبلا 5 ‘من ينتهي الكسر لتوليد 3 ‘واحد حبلا الحمض النووي (ssDNA) ذيول، وهي عملية يشار إليها باسم 5 ‘-3’ استئصال. مجمع MRN يبدأ استئصال نهاية الحمض النووي ويتم معالجة مزيد من استئصال في تركيبة مع BLM /EXO1 (بلوم متلازمة البروتين / exonuclease 1) أو BLM / DNA2 (الحمض النووي النسخ المتماثل ATP- تعتمد على helicase/nuclease)18,19,20,21,22. يتم تعزيز استئصال الحمض النووي نهاية بواسطة CtIP (CtBP التفاعل البروتين) من خلال تفاعلها المباشر مع MRN مجمع23 وتجنيد BRCA1 (سرطان الثدي نوع 1 البروتين القابلية للحساسية)24,25. بروتين النسخ المتماثل A (RPA) يربط على الفور إلى ssDNA يتعرض ومن ثم يتم إزاحة من قبل البروتين إعادة الكومبيناز RAD51 لتشكيل خيوط البروتين النووي الذي يحفز البحث المتجانسة وغزو حبلا26،27،28.
بدء عملية استئصال هو خطوة حاسمة لإصلاح المسار الاختيار. مرة واحدة وقد بدأت استئصال, ينتهي الحمض النووي تصبح ركائز الفقراء للربط بواسطة ku70/Ku80 heterodimer (مكون من مسار NHEJ) وتلتزم الخلايا إلى الموارد البشرية17,29,30. وKu70/Ku80 هيتيروديمر يربط إلى ينتهي DSB, تجنيد الحمض النووي-PKcs وp53 البروتين ملزمة 1 (53BP1)29,30. 53BP1 بمثابة حاجز للتشفط في G1، وبالتالي منع الموارد البشرية في حين تعزيز NHEJ31،32، ولكن تتم إزالته بطريقة BRCA1 تعتمد في المرحلة S، وبالتالي السماح للتشفط أن يحدث33،34. ولذلك، 53BP1 و BRCA1 تلعب أدوارا متعارضة في إصلاح DSB، مع 53BP1 كونه ميسر NHEJ في حين BRCA1 أعمال تمكين فواصل لإصلاح من خلال الموارد البشرية.
في المختبر، يمكن أن يكون سبب تكوين DSB الإشعاع المؤين (IR). في حين أن هذا المثال يستخدم جرعة عالية من 4 Gy، 1 غي و 2 غي أيضا إنشاء كمية كبيرة من DSBs، ومناسبة لتحليل تشكيل بؤر بواسطة البروتينات وفيرة. من المهم أن نلاحظ أن نوع وجرعة الإشعاع المستخدمة يمكن أن تؤدي إلى آفات مختلفة في الحمض النووي وفي الخلية: في حين أن الأشعة تحت الحمراء تحفز DSBs، يمكن أن تسبب أيضا فواصل حبلا واحد أو تعديل قاعدة (انظر35،,36 للاطلاع على مرجع على نقل الطاقة الخطي المشع (LET) ونوع الضرر الحمض النووي). لتحديد الحركية من المؤينة الناجمة عن الإشعاع تشكيل بؤر (IRIF) وإزالتها، والتي تشير إلى إصلاح الضرر وعكس من DDR تنشيط8،9،37،38، ويمكن رصد تشكيل بؤر في نقاط زمنية مختلفة بعد الإشعاع المؤين. ومن المعروف توقيت التنشيط وإزالة جميع البروتينات الضرر الحمض النووي الرئيسية39, ويتم التحقيق في العديد من علامات بديلة للأحداث الرئيسية. على سبيل المثال، pRPA، التي تمتلك تقارب عالية لSSDNA يستخدم كبديل للاستئصال كسر، يمكن استخدام البروتينات MRN (MRE11، RAD50، NBS1) والإفلات من الدهون لتقييم كفاءة استئصال أيضا. في حين RAD51، BRCA1، BRCA2 (سرطان الثدي نوع 2 البروتين القابلة للحساسية)، وPALB2 (شريك و localizer من BRCA2) يمكن رصدها لتقييم كفاءة الموارد البشرية، ووجود بروتينات كو أو 53BP1، وتستخدم كعلامات من NHEJ (الشكل 1).
كما البروتينات من آلات إصلاح الحمض النووي تجنيد بعضها البعض لكسر والتجمع في مجمعات فائقة، يمكن الاستدلال على التفاعلات الحمض النووي البروتين والبروتين من خلال اتباع التعريب الفردية مع مرور الوقت وتحليل التعريب المشترك للبروتينات، كما تصورها إشارات متداخلة في الخلية40،41،42. في خطوط الخلايا، يسمح إدخال طفرات نقطة أو حذف في جينات إصلاح الحمض النووي المحددة إما من خلال تحرير الجينوم، أو عن طريق التعبير الزائد عن مسوخ البلازميد، بالتحقيق في بقايا محددة ودورها المحتمل في التعرف على تلف الحمض النووي (على سبيل المثال، التعريب المشترك مع γH2AX) أو التجميع المعقد (التعريب المشترك مع بروتينات أخرى أو عدة)، فضلاً عن تأثيرها على إصلاح الحمض النووي. هنا، نستخدم الفلورا المناعية غير المباشرة كوسيلة للتحقيق في تشكيل وحل DSBs باتباع بؤر γH2AX مع مرور الوقت. كما نقدم مثالاً واحداً على تشكيل البؤر وتحليل التعريب المشترك من قبل لاعب رئيسي في إصلاح DSB: p53 ملزمة البروتين 1 (53BP1)32. كما ذكر سابقا، يعتبر 53BP1 مركزية لإصلاح الحمض النووي اختيار المسار. بعد تراكم 53BP1 وتوطينه المشترك مع γH2AX يوفر معلومات ثمينة عن مرحلة دورة الخلية ، وتراكم تلف الحمض النووي ، والمسار المستخدم لإصلاح DSBs. الغرض من تخصيص المناعة غير المباشرة هو تقييم كفاءة إصلاح تلف الحمض النووي في خطوط الخلايا ، بعد الأشعة تحت الحمراء كما في هذه الدراسة ، أو بعد التعرض لضغوط مختلفة في الخلية ، من ربط الحمض النووي إلى انسداد شوكة النسخ المتماثل (يتم توفير قائمة من العوامل الضارة للحمض النووي في الجدول 1).
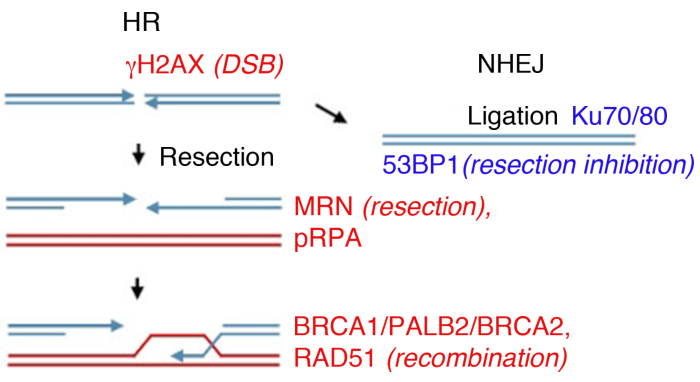
الشكل 1: فواصل الحمض النووي المزدوجة فواصل (DSB) مسارات الإصلاح.
إصلاح DSB ينطوي على مسارين رئيسيين: إعادة التركيب المتماثل (HR, اليسار) وغير متجانسة نهاية-صلة (NHEJ, حق). بعد الاستراحة، يتم تنشيط البروتينات للاحتفال بالكسر (γH2AX)، والمشاركة في استئصال نهاية (MRN)، أو معطف SsDNA (pRPA)، والترويج لإعادة الكومبونيشن (BRCA1، PALB2، BRCA2، RAD51) أو الحد من استئصال وترقية NHEJ (53BP1). البروتينات الأخرى المشاركة في إصلاح الضرر، ولكن البروتينات المذكورة تليها بشكل روتيني مناعي غير مباشر. الرجاء النقر هنا لعرض نسخة أكبر من هذا الرقم.
| عامل إتلاف الحمض النووي | آلية العمل | الجرعة الموصى بها |
| γ أشعة / الأشعة السينية | الاشعاع تشكيل فواصل مزدوجة تقطعت بها السبل مع بعض الآثار الخلوية غير المنضبط |
1-4 غي |
| 36 آر أيونات | الاشعاع تشكيل فواصل مزدوجة تقطعت بها السبل |
270 كيلو متر/ميكرومتر |
| α-الجسيمات | الاشعاع تشكيل فواصل مزدوجة تقطعت بها السبل |
116 كيلو/ميكرومتر |
| بلوميسين | مثبط تخليق الحمض النووي | 0.4-2 ميكروغرام/مل |
| كامبتويثسين | مثبط توبواomerase الأول | 10-200 nM |
| سيسبلاتين | وكيل Alkylating (تحريض الروابط المتقاطعة داخل العلامة التجارية) |
0.25-2 ميكرومتر |
| دوكسوروبسين | عامل المعايرة البينية مثبطات توبواomerase II |
10-200 nM |
| إيتوبوسيد | مثبطات توبواomerase II | 10 ميكرومتر |
| هيدروكسي يوريا | مثبط تخليق الحمض النووي (بواسطة ريبونوكليوتيد الاستئصال) |
10-200 ميكرومتر |
| ميثيل الميثان | وكيل Alkylating | 0.25-2 mM |
| ميتوميسين سي | وكيل Alkylating | 0.25-2 ميكرومتر |
| الأشعة فوق البنفسجية (UV) الخفيفة | تشكيل خافمات الثيميدين (توليد تشويه من سلسلة الحمض النووي) |
50-100 mJ / سم2 |
الجدول 1: العوامل السمية الجينية أمثلة على عوامل إتلاف الحمض النووي، وآلية عملها والأضرار الناجمة عن ذلك استناداً إلى التركيز المقترح للعمل.
