Le cellule umane sono costantemente esposte a una varietà di agenti dannosi per il DNA di varie origini. Le fonti esogene sono per lo più costituite da esposizione a radiazioni, sostanze chimiche (compresi agenti chemioterapici e alcuni antibiotici), e virus, mentre le principali fonti endogene includono errori nella replicazione del DNA e nello stress ossidativo. Gli effetti diretti dell’esposizione genotossica possono variare da una base modificata a una rottura del doppio filamento del DNA potenzialmente letale (DSB), a seconda dello stress e della dose di esposizione. In definitiva, danni al DNA non riparati o riparati in modo improprio possono portare all’accumulo di mutazioni, riarrangiamenti genomici, instabilità del genoma e alla fine portare alla carcinogenesi1. Le cellule dei mammiferi hanno sviluppato percorsi complessi per riconoscere specifici tipi di danni al DNA2,3 e ripararli in modo tempestivo, sincronizzati con la progressione del ciclo cellulare.
Le radiazioni ionizzanti (IR) danneggiano la doppia elica del DNA e creano rotture a doppio filamento (DSB), una delle forme più deleteri di danno al DNA. Il complesso MRN (MRE11, RAD50, NBS1) funziona come un sensore di DNA termina e attiva la proteina chinasi ataxia telangiectasia mutato (ATM)4,5. Dopo l’attivazione iniziale di ATM da fini dna, ATM innesca una cascata di eventi DDR sul sito della rottura, avviando con un evento chiave, la fosforilazione della variante istone H2AX6. La fosforilazione H2AX sui residui S139 la attiva in γH2AX, che si estende su regioni fino a megabase intorno alla lesione del DNA6,7,8,9. Questo evento aumenta l’accessibilità del DNA, portando al reclutamento e all’accumulo di altre proteine di riparazione del DNA7. Poiché γH2AX è abbondantemente e specificamente indotto DSBs circostanti, può essere facilmente visualizzata utilizzando anticorpi specifici, ed è comunemente usato come marcatore surrogato per DSB nel campo di riparazione del DNA. Una volta segnalata la rottura, le cellule attivano le loro vie di riparazione del DNA ed elaborano il danno al DNA. La proteina MDC1 (mediatore della proteina checkpoint del danno al DNA 1) lega direttamente γH2AX10,interagisce con ATM11 e anche con NBS112,,13. Contribuisce ad aumentare la concentrazione del complesso MRN presso il DSB e ad avviare un ciclo di feedback ATM positivo. γH2AX viene rapidamente rimosso una volta riparata la rottura, consentendo di conseguenza il monitoraggio dell’autorizzazione DSB. Seguita dalla microscopia, la diminuzione della γH2AX nel tempo fornisce una misurazione indiretta delle rotture residue e dell’efficienza di riparazione del DNA.
Le cellule eucariotiche possono riparare i DSB da diversi percorsi, mentre le due principali sono l’unione finale non omologa (NHEJ) e la ricombinazione omologa (HR) (Figura 1). NHEJ lega essenzialmente il DNA a doppio filamento termina senza l’uso di omologia estesa e opera per tutto il ciclocellulare 14,15. HR diventa predominante durante le fasi S e G2, ed è altrimenti represso, dal momento che richiede una sorella cromata come modello omologa per lariparazione 14,16. La scelta del percorso tra NHEJ e HR dipende non solo dalla vicinanza fisica della sorella cromatica, ma anche dall’estensione della resezione finale del DNA17, che inibisce NHEJ.
La riparazione DSB dipendente dall’omologia inizia con la degradazione nucleolitica del filamento 5′ dalle estremità di rottura per generare code di DNA a 3′ filamento singolo (ssDNA), un processo denominato resezione 5′-3′. Il complesso MRN avvia la resezione finale del DNA e l’ulteriore resezione viene elaborata in combinazione con BLM/EXO1 (Proteina sindrome di Bloom/esonucleasi 1) o BLM/DNA2 (replicazione del DNA Elicase/nucleasi dipendente da ATP)18,19,20,21,22. La resezione finale del DNA è potenziata dal CtIP (proteina che interagisce con il CtBP) attraverso la sua interazione diretta con il complesso MRN23 e il reclutamento di BRCA1 (proteina di suscettibilità al tipo 1 del cancro al seno)24,25. La proteina di replica A (RPA) si lega prontamente alla ssDNA esposta e viene quindi spostata dalla proteina ricombinata RAD51 per formare un filamento nucleoproteina che catalizza la ricerca omologa e l’invasionedel filamento 26,27,28.
L’avvio della resezione è un passo critico per la scelta del percorso di riparazione. Una volta avviata la resezione, le estremità del DNA diventano substrati poveri per la legazione da parte dell’eterodimero Ku70/Ku80 (componente del percorso NHEJ) e le cellule sono impegnate in HR17,29,30. L’eterodimero Ku70/Ku80 si lega alle estremità DSB, reclutando DNA-PKcs e p53 Binding Protein 1 (53BP1)29,30. 53BP1 agisce come una barriera alla resezione in G1, bloccando così le risorse umane durante la promozione di NHEJ31,32, ma viene rimosso in modo dipendente da BRCA1 nella fase S, consentendo di conseguenza la resezione33,34. Pertanto, 53BP1 e BRCA1 svolgono ruoli opposti nella riparazione DSB, con 53BP1 essendo un facilitatore NHEJ mentre BRCA1 agisce consentendo interruzioni per la riparazione attraverso HR.
In laboratorio, la formazione di DSB può essere indotta da radiazioni ionizzanti (IR). Mentre questo esempio utilizza una dose elevata di 4 Gy, 1 Gy e 2 Gy anche creare una quantità significativa di DSB, adatto per l’analisi della formazione di foci da proteine abbondanti. È importante notare che il tipo e la dose di radiazioni utilizzate possono portare a diverse lesioni nel DNA e nella cellula: mentre l’IR induce ISB, può anche causare rotture di singoli filamenti o modificazione della base (vedi35,36 per un riferimento sul trasferimento di energia lineare di irradiazione (LET) e sul tipo di danno al DNA). Per determinare la cinetica della formazione di foci ionizzanti indotta da radiazioni (IRIF) e la loro distanza, che indicano la riparazione del danno e l’inversione del DDRattivato 8,9,37,38, la formazione di foci può essere monitorata in diversi punti di tempo dopo la radiazione ionizzante. La tempistica di attivazione e di sgombero di tutte le principali proteine danno del DNAè nota 39, e molti sono studiati come marcatori surrogati di eventi chiave. Ad esempio, pRPA, che possiede un’elevata affinità per ssDNA viene utilizzato come surrogato della resezione di rottura, le proteine MRN (MRE11, RAD50, NBS1) e le esonucleasi possono essere utilizzate anche per valutare l’efficienza della resezione. Mentre RAD51, BRCA1, BRCA2 (proteina di suscettibilità al cancro al seno 2) e PALB2 (partner e localizzatore di BRCA2) possono essere monitorati per valutare l’efficienza delle risorse umane, la presenza delle proteine Ku o 53BP1, vengono utilizzati come marcatori di NHEJ(Figura 1).
Poiché le proteine del macchinario per la riparazione del DNA si reclutano a vicenda per la rottura e l’assemblaggio in super-complessi, le interazioni DNA-proteina e proteina-proteina possono essere dedotte seguendo la loro localizzazione individuale nel tempo e analizzando la co-localizzazione delle proteine, come viene visualizzazioneto da segnali sovrapposti nella cella40,41,42. Nelle linee cellulari, l’introduzione di mutazioni puntino o la cancellazione in specifici geni di riparazione del DNA sia attraverso l’editing del genoma, sia per sovraespressione di mutanti a base di plasmidi, consente di eseguire lo studio di residui specifici e del loro possibile ruolo nel riconoscimento del danno al DNA (ad esempio, la co-localizzazione con γH2AX) o di un assemblaggio complesso (co-localizzazione con un’altra o più proteine), nonché il loro impatto sulla riparazione del DNA. Qui, usiamo l’immunofluorescenza indiretta come mezzo per studiare la formazione e la risoluzione dei DSB seguendo γH2AX foci nel tempo. Presentiamo anche un esempio di formazione di foci e analisi di co-localizzazione da parte di un attore importante nella riparazione DSB: p53 Binding Protein 1 (53BP1)32. Come accennato in precedenza, 53BP1 è considerato centrale per la scelta del percorso di riparazione del DNA. Dopo l’accumulo di 53BP1 e la sua co-localizzazione con γH2AX fornisce preziose informazioni sulla fase del ciclo cellulare, l’accumulo di danni al DNA e il percorso utilizzato per riparare i DSB. Lo scopo dell’immunolocalizzazione indiretta è quello di valutare l’efficienza della riparazione dei danni al DNA nelle linee cellulari, seguendo IR come in questo studio, o dopo l’esposizione a varie sollecitazioni nelle cellule, dal collegamento incrociato del DNA al blocco della forchetta di replica (un elenco di agenti dannosi per il DNA è fornito nella tabella 1).
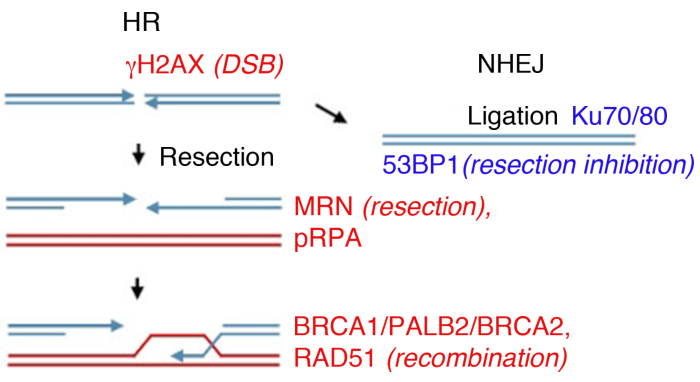
Figura 1: Vie di riparazione del doppio filamento del DNA (DSB).
La riparazione DSB prevede due percorsi principali: la ricombinazione omologa (HR, a sinistra) e l’end-joining non omologho (NHEJ, a destra). Dopo la pausa, le proteine si attivano per contrassegnare la rottura (γH2AX), partecipare alla resezione finale (MRN), rivestire la ssDNA resected (pRPA), promuovere la ricombinazione (BRCA1, PALB2, BRCA2, RAD51) o limitare la resezione e promuovere NHEJ (53BP1). Altre proteine partecipano alla riparazione dei danni, ma le proteine elencate sono regolarmente seguite dall’immunofluorescenza indiretta. Si prega di fare clic qui per visualizzare una versione più grande di questa figura.
| Agente dannoso del DNA | Meccanismo d’azione | Dose raccomandata |
| γ/raggi X | Radiazione Formazione di rotture a doppio filamento con alcuni effetti cellulari incontrollati |
1-4 Gy |
| Il numero 36 del 36 Ar ioni | Radiazione Formazione di rotture a doppio filamento |
270 keV/m |
| particelle z | Radiazione Formazione di rotture a doppio filamento |
116 keV/m |
| Bleomicina | Inibitore della sintesi del DNA | 0,4-2 g/mL |
| Camptothecin | Inibitore della topoisomerasi I | 10-200 nM |
| Cisplatino | Agente di Alkylating (inducendo collegamenti incrociati intrastranti) |
0,25-2 M |
| Doxorubicina | Agente intercalante Inibitore della topoisomera II |
10-200 nM |
| Etoposide | Inibitore della topoisomera II | 10 SM |
| Hydroxyurea | Inibitore della sintesi del DNA (per ribonucleotide reductase) |
10-200 M |
| Metano metilesulfonato | Agente di Alkylating | 0,25-2 mM |
| Mitomicina C | Agente di Alkylating | 0,25-2 M |
| Luce ultravioletta (UV) | Formazione di dimeri di timodina (generando distorsione della catena del DNA) |
50-100 mJ/cm2 |
Tabella 1: agenti genotossici. Esempi di agenti dannosi per il DNA, il loro meccanismo d’azione e il danno indotto sulla base della concentrazione di lavoro suggerita.
