İnsan hücreleri sürekli çeşitli kökenlerden DNA zararlı ajanlar çeşitli maruz kalırlar. Ekzojen kaynaklar çoğunlukla radyasyona maruz ilerler, kimyasallar (kemoterapötik ajanlar ve bazı antibiyotikler dahil) ve virüsler, ana endojen kaynaklar DNA replikasyonu ve oksidatif stres hataları içerir iken. Genotoksik maruziyetin doğrudan etkileri, strese ve maruz kalma dozuna bağlı olarak modifiye edilmiş bir bazdan potansiyel olarak öldürücü DNA çift iplikçik kırılamı (DSB) arasında değişebilir. Sonuçta, onarılmamış veya yanlış onarılmış DNA hasarı mutasyonların birikimine yol açabilir, genomik yeniden düzenlemeler, genom istikrarsızlık ve sonunda karsinogenez yol 1 .1 Memeli hücreleri DNA hasarı belirli türleri tanımak için karmaşık yollar gelişti2,3 ve zamanında onları onarmak, hücre döngüsü ilerlemesi ile senkronize.
İzole radyasyon (IR) DNA çift sarkıklık zarar ve çift iplikçik sonları oluşturur (DSBs), DNA hasarının en zararlı formlarından biri. MRN (MRE11, RAD50, NBS1) DNA sensörü olarak karmaşık fonksiyonları biter ve protein kiaz ataksi telanjiektazi mutasyona (ATM)4,5aktive . DNA biter tarafından ATM ilk aktivasyonu ardından, ATM önemli bir olay ile başlayan, mola yerinde DDR olaylar bir çağlayan tetikler, histon varyantı H2AXfosforilasyon 6. Kalıntı S139 üzerindeki H2AX fosforilasyon γH2AX içine aktive, DNA lezyonu etrafında megabazlar kadar bölgeleri kapsayan6,7,8,9. Bu olay DNA erişilebilirliğini artırır, işe alma ve diğer DNA onarım proteinleri birikimine yol 7. γH2AX bol ve özellikle DSB’leri çevreleyen indüklenen olduğundan, belirli antikorlar kullanılarak kolayca görselleştirilebilir ve genellikle DNA onarım alanında DSB’ler için bir taşıyıcı belirteç olarak kullanılır. Mola sinyali verildikten sonra hücreler DNA tamir yollarını aktive eder ve DNA hasarını işlerler. Protein MDC1 (DNA hasar kontrol noktası protein arabulucusu 1) doğrudan γH2AX10bağlanır , ATM11 ile etkileşime ve ayrıca NBS112,13. DSB’deki MRN kompleksinin konsantrasyonunun artırılmasına ve olumlu bir ATM geri besleme döngüsübaşlatılmasına katkıda bulunur. γH2AX, mola onarıldıktan sonra hızla çıkarılır ve dsb açıklığının izlenmesine izin verilir. Mikroskopinin ardından γH2AX’ın zaman içinde azalması, artık sonları ve DNA onarım veriminin dolaylı olarak ölçülmesini sağlar.
Ökaryotik hücreler DSB’leri çeşitli yollarla onarabilirler, iki ana hücre homolog olmayan son birleştirme (NHEJ) ve homolog rekombinasyon (HR) dir. NHEJ aslında genişletilmiş homoloji kullanmadan DNA çift iplikçikli biter ligates ve hücre döngüsü boyunca çalışır14,15. Hr S ve G2 aşamalarında baskın hale gelir, ve aksi takdirde bastırılmış, onarım için homolog bir şablon olarak bir kardeş kromatit gerektirir beri14,16. NHEJ ve İk arasındaki yol seçimi sadece kardeş kromatit fiziksel yakınlığına bağlıdır, ama aynı zamanda DNA sonu rezeksiyon genişletmekbağlıdır 17, Hangi NHEJ inhibe.
Homolojiye bağımlı DSB onarımı, 5′ ipliğin kopma uçlarından nükleolitik bozulma ile başlatılarak 3′ tek iplikçikli DNA (ssDNA) kuyrukları oluşturmak için, 5′-3′ rezeksiyon olarak adlandırılan bir süreçtir. MRN kompleksi DNA sonu rezeksiyonu başlatır ve daha fazla rezeksiyon BLM / EXO1 (Bloom sendromu protein / ekonenaz 1) veya BLM / DNA2 (DNA replikasyonu ATP bağımlı helicase / nükleaz)18,19,20,21,22ile birlikte işlenir . DNA sonu rezeksiyonu CTIP (CtBP-etkileşen protein) mrn kompleksi ile doğrudan etkileşim yoluyla geliştirilmiştir23 ve BRCA1 işe (meme kanseri tip 1 duyarlılık proteini)24,25. Replikasyon proteinI A (RPA) derhal açığa çıkan ssDNA bağlanır ve daha sonra homolog arama ve iplikçik invazyonu katalizler bir nükleoprotein filament oluşturmak için rekombinaz protein RAD51 tarafından yerinden edilir26,27,28.
Rezeksiyon un başlatılması onarım yolu seçimi için kritik bir adımdır. Bir kez rezeksiyon başladı, DNA biter Ku70/Ku80 heterodimer (NHEJ yolu bileşeni) tarafından bağlayıcı için zayıf substratlar haline gelir ve hücreler HR17,,29,30kararlıdır . Ku70/Ku80 heterodimer DSB biter bağlanır, DNA-PKcs ve p53 Bağlayıcı Protein 1 (53BP1)29,,30işe. 53BP1 G1 rezeksiyon için bir bariyer olarak görür, böylece NHEJ teşvik ederken İk engelleme31,32, ama S fazında brca1 bağımlı bir şekilde kaldırılır, sonuç olarak rezeksiyon oluşmasına izin33,34. Bu nedenle, 53BP1 ve BRCA1 DSB onarımında karşıt roller oynarken, 53BP1 NHEJ kolaylaştırıcısı olurken BRCA1, hr aracılığıyla onarım molalarını sağlar.
Laboratuvarda, DSB oluşumu iyonlaştırıcı radyasyon (IR) ile indüklenebilir. Bu örnekte yüksek dozda 4 Gy, 1 Gy ve 2 Gy de bol proteinlerle fosi oluşumunun analizi için uygun miktarda DSB oluşturur. Kullanılan radyasyonun türü ve dozu DNA’da ve hücrede farklı lezyonlara yol açabilir dikkat etmek önemlidir: IR DSB’ler indükler iken, aynı zamanda tek iplikçik kırıkları veya baz modifikasyonu neden olabilir (ışınlama lineer enerji transferi bir referans için35,36 bakınız (LET) ve DNA hasarı türü). Radyasyona bağlı fok oluşumunun iyonlaştırıcı kinetikbelirlemek için (IRIF) oluşumu ve bunların temizliği, aktif DDR8,9,,37,38hasar ve ters onarım gösteren, fosi oluşumu iyonlaştırıcı radyasyon sonra farklı zaman noktalarında izlenebilir. Aktivasyon zamanlaması ve tüm büyük DNA hasarı proteinlerin temizlenmesi39bilinmektedir , ve birçok önemli olayların vekil işaretleri olarak araştırılmaktadır. Örneğin, ssDNA’ya yüksek afiniteye sahip olan pRPA, kırılma rezeksiyonunun taşıyıcı sı, MRN proteinleri (MRE11, RAD50, NBS1) ve eksonüekleaseler rezeksiyon verimliliğini değerlendirmek için de kullanılabilir. RAD51, BRCA1, BRCA2 (meme kanseri tip 2 duyarlılık proteini) ve PALB2 (BRCA2’nin partneri ve lokalizatörü) HR verimliliğini değerlendirmek için izlenebilse de, Ku proteinlerinin varlığı veya 53BP1, NHEJ’nin belirteçleri olarak kullanılır (Şekil 1).
DNA onarım makinelerinin proteinleri birbirlerini süper komplekslerde biraraya getirerek biraraya getirdikçe, DNA-protein ve protein-protein etkileşimleri zaman içinde bireysel lokalizasyonlarını takip ederek ve proteinlerin eş lokalizasyonunu analiz ederek çıkarılabilir, hücre40,41,42’dekisinyalleri örtüşerek görselleştirilmiştir. Hücre hatlarında, belirli DNA onarım genlerinde nokta mutasyonlarının veya silmenin genom düzenlemesi yoluyla ya da plazmid tabanlı mutantların aşırı ekspresyonu yoluyla başlatılması, spesifik kalıntıların araştırılmasına ve DNA hasarının tanınmasındaki olası rollerinin (örneğin, γH2AX ile birlikte lokalizasyon) veya karmaşık montajın (başka bir veya birkaç proteinle birlikte lokalizasyon) yanı sıra DNA onarımı üzerindeki etkilerinin araştırılmasına olanak sağlar. Burada, dsbs oluşumunu ve çözünürlüğünü araştırmak için bir ortalama olarak dolaylı immünfloresans kullanın γH2AX foci zaman içinde takip ederek. Ayrıca DSB onarımönemli bir oyuncu tarafından foci oluşumu ve co-lokalizasyon analizi bir örnek satıyoruz: p53 Bağlayıcı Protein 1 (53BP1)32. Daha önce de belirtildiği gibi, 53BP1 DNA onarım yolu seçimi merkezi olarak kabul edilir. ΓH2AX ile birlikte 53BP1 birikimi ve birlikte lokalizasyonu hücre döngüsü fazı, DNA hasar birikimi ve DSB’leri onarmak için kullanılan yol hakkında değerli bilgiler sağlar. Dolaylı immünolokalizasyonun amacı, bu çalışmada olduğu gibi IR’yi takip ederek veya hücredeki çeşitli streslere maruz kaldıktan sonra, DNA çapraz bağlantısından çoğaltma çatalının tıkanması için hücre hatlarındaki DNA hasar onarımının etkinliğini değerlendirmektir (DNAzararlı ajanların listesi Tablo 1’de verilmiştir).
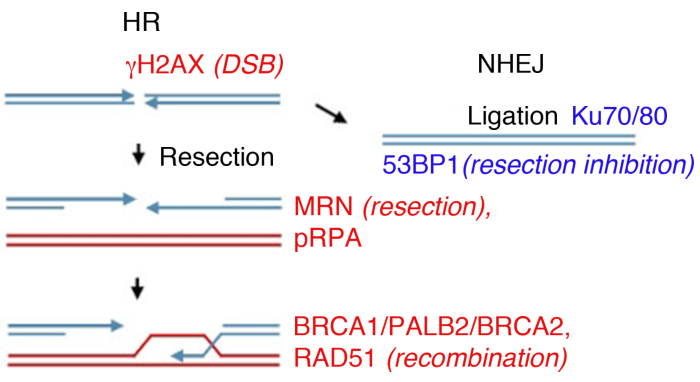
Şekil 1: DNA çift iplikçik kırıkları (DSB) onarım yolları.
DSB onarımı iki ana yolu içerir: Homolog Rekombinasyon (İk, sol) ve Non-Homolog End-Joining (NHEJ, sağ). Aradan sonra proteinler break (γH2AX) işaretlemek için aktive olsun, son rezeksiyon (MRN) katılmak, rezeke ssDNA (pRPA), rekombinasyon teşvik (BRCA1, PALB2, BRCA2, RAD51) veya rezeksiyon sınırlamak ve NHEJ (53BP1) teşvik. Diğer proteinler hasar onarımına katılırlar, ancak listelenen proteinler rutin olarak dolaylı immünfloresans tarafından takip edilir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
| DNA zararlı madde | Etki mekanizması | Önerilen doz |
| γ-ışınları/X-ışınları | Radyasyon Bazı kontrolsüz hücresel etkilerle çift iplikli kırıkların oluşması |
1-4 Gy |
| 36.000 Ar iyonları | Radyasyon Çift iplikli sonları oluşumu |
270 keV/μm |
| α parçacıkları | Radyasyon Çift iplikli sonları oluşumu |
116 keV/μm |
| Bleomisin | DNA sentezinin inhibitörü | 0.4-2 μg/mL |
| Camptothecin | Topoisomerase Inhibitörü I | 10-200 nM |
| Sisplatin | Alkylating ajan (intrastrand crosslinks indükleyen) |
0,25-2 μM |
| Doksorubisin | Ara ajan Topoisomerase Inhibitörü II |
10-200 nM |
| Etoposide | Topoisomerase Inhibitörü II | 10 μM |
| Hidroksyurea | DNA sentezinin inhibitörü (ribonükleotid redüktaz tarafından) |
10-200 μM |
| Metil metansülfonat | Alkylating ajan | 0,25-2 mM |
| Mitomisin C | Alkylating ajan | 0,25-2 μM |
| Ultraviyole (UV) ışık | Timidin dimers oluşumu (DNA zincirinin bozulmasını üreten) |
50-100 mJ/cm2 |
Tablo 1: Genotoksik ajanlar. DNA’ya zarar veren ajanlar, etki mekanizmaları ve önerilen çalışma konsantrasyonuna dayalı hasar örnekleri.
