Rumtemperatur (RT) makromolekylær krystallografi er populær igen inden for strukturbiologi samfund. Udviklingen af røntgenfri elektronlaser (XFEL) lyskilder har ansporet udviklingen af RT-prøveleveringsmetoder 1,2,3,4, og disse metoder er nu blevet anvendt på synkrotroner 5,6,7,8. RT-metoder åbner ikke kun muligheden for pumpesondeeksperimentelle strageties 9,10,11,12, men der er også stigende beviser for, at de fremmer alternative konformationstilstande inden for proteiner 13,14,15,16,17.
Hovedårsagen til, at kryo-metoder fik trækkraft over RT-tilgange i slutningen af 1990’erne, var imidlertid opbremsningen af strålingsskader ved krystaltemperaturerunder nul 18. Kryo-metoder19 begyndte at muliggøre indsamling af et komplet datasæt fra en enkelt proteinkrystal. Moderne RT-metoder ved XFELs og synkrotroner løste problemet med enkeltkrystalstrålingsskader ved udvikling af hurtige (> 100 Hz) krystalleveringsstrategier 1,2,3,4. Disse metoder giver mulighed for indsamling af et komplet datasæt fra tusindvis af individuelt eksponerede krystaller. Disse RT-leveringsmetoder kræver derfor produktion af store mængder opløsninger indeholdende homogene mikrokrystaller (> 100 μL < 50 μm krystaller). Men da kryometoder har tendens til kun at kræve enkeltkrystaller, er metoder til at skabe sådanne mikrokrystallinske opslæmninger i øjeblikket ikke allestedsnærværende på tværs af proteinkrystallografilaboratorier.
Der er eksempler i litteraturen på, hvordan man gør dele af mikrokrystallisationsoptimeringsproceduren for serielle krystallografiprøver. Her skal der skelnes mellem membran og opløselige proteiner. Protokoller til optimering af væksten af mikromembranproteinkrystaller dyrket i monoolein (eller et andet lipid) til lipidisk kubisk fase (LCP) er blevet godt beskrevet20,21,22. Imidlertid mangler der generelt metoder til mikrokrystallisation af opløselige proteiner, herunder membranproteiner dyrket under ikke-LCP-betingelser. Tidligere undersøgelser har fokuseret på specifikke dele af processen, såsom mikrokrystalscreening 23,24, forbedring af kimdannelse24 og skalering ved hjælp af fri grænsefladediffusion 25, men ikke en komplet metode.
Imidlertid blev der for nylig beskreveten metode 26 , der forsøger at tilbyde en komplet protokol. Ligesom mange aspekter af proteinkrystallografi er det ikke nyt. Mange af de foreslåede ideer blev allerede beskrevet af Rayment (2002)27. Metoden har til formål at vise krystallografer, hvordan man udfører konverteringen fra en enkelt krystal dyrket ved hjælp af dampdiffusion til en batchmetode til dyrkning af tusindvis af mikrokrystaller. Metoden fokuserer på dampdiffusion som et fælles udgangspunkt, da 95% af alle Protein Data Bank (PDB) aflejringer kommer fra krystaller dyrket i dampdiffusionsplader26. Dampdiffusion er imidlertid ikke den ideelle metode til mikrokrystallisation26, så der beskrives en metode til at konvertere dampdiffusion til batchkrystallisation. Når krystaller kan dyrkes i batch, bliver skaleringsruter til større mængder mere praktiske. I betragtning af proteinkrystalliseringens luner vil forfatterne understrege, at denne metode ikke er fejlsikker. Protokollen skal dog i det mindste give et indblik i et proteins ‘krystalliseringsrum’.
Denne metode er afhængig af proteinkrystalliseringsfasediagrammet, og hvordan en forståelse af dette diagram kan fungere som vejledning under mikrokrystallisationsoptimering. Et proteinfasediagram er almindeligvis afbildet som et x/y-plot med udfældnings- og proteinkoncentrationer på henholdsvis x- og y-akserne (figur 1A). Fra punktet rent vand (nederste venstre hjørne – figur 1A) øges koncentrationen af både protein og bundfald, indtil opløselighedslinjen er nået. Opløselighedslinjen markerer overmætningspunktet (lilla linje – figur 1A). Når et protein er overmættet, bliver opløsningen termodynamisk ustabil og begynder at adskille sig i to faser: ‘proteinrig’ og en stabil mættet opløsning. Denne adskillelse kan forekomme hvor som helst uden for opløselighedslinjen, og dens kinetik afhænger af proteinets egenskaber og opløsningens komponenter.
Når protein- og bundfaldkoncentrationerne er for store, nedbrydes proteinet ustabilt ud af opløsningen og resulterer i amorft bundfald (lyserødt område – figur 1A). Imidlertid kan ordnet faseadskillelse forekomme i kimdannelsesområdet [se Garcia-Ruiz (2003) 28 for detaljeret beskrivelse], og krystalnukleanter har tilbøjelighed til at danne (grøn region – figur 1A). Nukleation og vækst fjerner protein fra opløsningen og flytter dråben ind i det metastabile område, hvor væksten kan fortsætte, indtil opløselighedslinjen er nået [se McPherson og Kuznetsov (2014) 29 for detaljeret diskussion]. Diagrammet er for langt de fleste krystalliseringsbetingelser en grov oversimplificering30. Uanset dette er diagrammet dog stadig af stor nytte for mikrokrystallografer, da kortlægningen af diagrammet gør det muligt at bestemme opløselighedslinjen og kinetikken for kimdannelse.
Med hensyn til at skabe mikrokrystaller er de to faktorer under krystallisering, der skal optimeres, antallet af krystaller (Xn) og deres gennemsnitlige, længste dimension (Xs). X n vil være proportional med antallet af kimdannelseshændelser (n ) (Eq. 1).
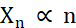 Eq. 1
Eq. 1
X s er proportional med koncentrationen af frit protein over opløselighedslinjen (Ps) divideret med Xn (Eq. 2).
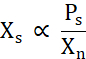 Eq. 2
Eq. 2
I en perfekt situation ville enhver kimdannelsesbegivenhed give en mulig krystal, og hver eneste af disse krystaller ville have lige adgang til det tilgængelige protein i opløsning. Figur 2 er en grafisk repræsentation fra et ideelt scenario af forholdet mellem Xn og Xs. I praksis er den vigtigste kontrol, en krystallograf har over Xn og Xs , ved at påvirke mængden af kimdannelse eller ved tilsætning af frøkrystaller. Mikrokrystallografen skal bedømme, hvordan man øger Xn , således at der både kan skabes en passende krystalkoncentration og krystalstørrelse.
De fleste krystalliseringsteknikker kræver en “overgangsperiode” (figur 1B). For eksempel i et dampdiffusionseksperiment, ved blanding af protein- og bundfældningsopløsningerne, vil koncentrationerne af hver ændre sig, når dråben udligner sig med brøndopløsningen. Man håber, at disse ændringer gradvist vil overføre faldet til kimdannelseszonen, hvor tilbøjeligheden til krystallisation vil stige. Når krystaller begynder at nukleere og vokse, vil mængden af protein i opløsning begynde at falde, hvilket reducerer sandsynligheden for yderligere kimdannelse. Den ultimative mængde kimdannelse vil være protein- og tilstandsspecifik og også afhængig af dybden af penetration af i nukleationszonen. I betragtning af den begrænsede kimdannelseszonepenetration af metoder, der kræver et overgangstrin, vil kimdannelsesniveauet i sidste ende være begrænset til kimdannelseshastigheden ved metastabil-nukleationsregionsgrænsen.
På grund af vigtigheden af at kunne forbedre kimdannelsesniveauet for en mikrokrystallograf er det vigtigt at flytte til en batchkrystallisationsmetode. Batch kan drage større fordel af hele nukleationsområdet (figur 1C). I batchmetoder er ideen at blande proteinet og bundfaldet sammen, således at der skabes en overmættet opløsning uden behov for ændringer i komponentkoncentrationer. Nukleation bør være mulig umiddelbart efter blanding. Batchmetoder tillader derfor, at hele kimdannelseszonen teoretisk nås. Enhver stigning i kimdannelseskinetik ud over metastabil-nukleationsgrænsen kan derefter udnyttes.
Hvis det basale niveau af krystalkimdannelse ikke er nok til at generere et stort Xn, kan mikrosåningsmetoder anvendes. Ved mikrosåning brydes forvoksede krystaller op for at skabe en opslæmning af krystallinske fragmenter, der kan fungere som et stillads for frisk krystalvækst31,32. Mikrosåning har været meget udbredt i seriel krystallografisk prøveforberedelse som en måde at øge Xn uden behov for at øge krystalkimdannelsen (figur 1C).
Overgangen fra dampdiffusion til batch kan visualiseres på et fasediagram som at flytte det eksperimentelle udgangspunkt fra enten de ikke-overmættede eller metastabile regioner til kimdannelseszonen. Dette kan gøres ved at øge protein- og/eller udfældningskoncentrationerne og/eller forholdet mellem de to inden for faldet (figur 1D) og observere, hvilke betingelser der giver krystaller, der viser sig hurtigt (< 24 timer)26. Fuldstændig dampdiffusionsdråbeligevægt kan tage dage eller uger33. Derfor, ved at lede efter betingelser, der viser hurtigt fremkomne krystaller, kan batchbetingelser findes uden at skulle flytte til alternative krystalliseringsscreeningsformater såsom mikrobatch34,35,36,37.
Når kimdannelseszonen er fundet, er der fundet en batchtilstand, og der kan oprettes et morfogram – her et groft fasediagram. Morfogrammet er af stor nytte, når man overvejer, om man skal bruge en podet batch eller lige batchprotokol. Ved at plotte Xn som funktion af protein- og bundfældningskoncentrationen kan der foretages en vurdering af kimdannelseskinetikken26. Hvis X n forbliver lav over hele kimdannelsesområdet, kan det være nødvendigt med frøbatch for at gøre Xn stor nok til at begrænse krystalvækst. Denne vurdering er det første skridt i processen med at skalere til større mængder (> 100 μL).
Denne metode blev designet således, at den kunne udføres i de fleste krystallisationslaboratorier ved hjælp af standard dampdiffusionskrystallisationsudstyr. Der er også udført mange undersøgelser, som beskriver teknikker til at lette mange dele af denne proces, hvis udstyret er tilgængeligt. Disse omfatter, men er ikke begrænset til, dynamisk lysspredning (DLS) 25,27, ikke-lineær billeddannelse 20,24,25, pulverdiffraktion 20,24,27 og elektronmikroskopi 26 [se Cheng et al. (2020) 40 for en god gennemgang].
Formålet med dette arbejde er at give en visuel demonstration af metoden til overgang fra lille volumen ( 100 μL) batchkrystallisering. Endothiapepsin fra Cryphonectria parasitica er blevet brugt som et eksempelsystem til demonstration af denne oversættelse. Den type eksperiment og prøveleveringsmetode, som mikrokrystallerne er nødvendige for, vil påvirke det ideelleX-s output26. For blandingsforsøg, der kræver en millisekundtidsopløsning41 eller gasdynamiske virtuelle dyser42, kan en endelig Xs på < 5 μm være ønskelig. I dette tilfælde var målet at producere proteinkrystaller, der diffrakterer til ca. 1,5 Å, til et fotonaktiveret pumpesondeeksperiment og ved hjælp af en leveringsmetode med fast mål.
For at give en illustration af prøvekravene til et sådant serielt krystallografieksperiment ved anvendelse af endothiapepsin viser tabel 1 de eksperimentelle parametre for et hypotetisk eksperiment. Eksempeloplysningerne var baseret på den protokol, der er beskrevet nedenfor. I betragtning af nogle konservative skøn over hitrater og dataindsamlingskrav er 50 mg det samlede prøveforbrugsestimat for hele eksperimentet.
Figur 3 viser et rutediagram over den komplette optimeringsproces fra indledende dampdiffusionskrystallisation i lille volumen til storskalabatch. For de fleste serielle krystallografiprojekter begynder denne protokol på trin 2: ‘overgang til batch’, da målproteinet allerede vil være krystalliseret. Trin 1 er dog medtaget for fuldstændighed og for at minde læserne om dens betydning. At finde en tilstand, der giver anledning til en godt diffrakterende, enkelt, stor krystal, er det bedste udgangspunkt for mikrokrystaloptimering. I trin 2 kan denne tilstand derefter optimeres fra dampdiffusion til batch, og et morfogram af nukleations- og metastabile regioner kan plottes. Når dette er gjort, kan batchbetingelsen skaleres til større mængder i trin 3. Ved afslutningen af rutediagrammet vil en krystallograf have skabt en gentagelig, stor volumen (> 100 μL), mikrokrystallisation, batchprotokol for endothiapepsin. Denne metode kan derefter anvendes på deres særlige protein af interesse.
