DNA-databehandling bruker et sett med designet oligonukleotider som medium for beregning. Disse oligonukleotidene er programmert med sekvenser for dynamisk montering i henhold til brukerspesifikk logikk og reagerer på spesifikke nukleinsyreinnganger. I konseptgodkjenningsstudier består utgangen av beregningen vanligvis av et sett fluorescerende merket oligonukleotider som kan oppdages via gelelektroforese eller fluorescensplatelesere. I løpet av de siste 30 årene har stadig mer komplekse DNA-beregningskretser blitt demonstrert, for eksempel ulike digitale logikkkaskader, kjemiske reaksjonsnettverk og nevrale nettverk1,2,3. For å hjelpe til med utarbeidelsen av disse DNA-kretsene, har matematiske modeller blitt brukt til å forutsi funksjonaliteten til syntetiske genkretser4,5og beregningsverktøy er utviklet for ortogonal DNA-sekvensdesign6,7,8,9,10 . Sammenlignet med silisiumbaserte datamaskiner inkluderer fordelene med DNA-datamaskiner deres evne til å kommunisere direkte med biomolekyler, operere i løsning i fravær av strømforsyning, samt deres generelle kompaktitet og stabilitet. Med ankomsten av neste generasjons sekvensering har kostnadene ved å syntetisere DNA-datamaskiner gått ned de siste to tiårene med en hastighet raskere enn Moores lov11. Anvendelser av slike DNA-baserte datamaskiner begynner nå å dukke opp, for eksempel for sykdomsdiagnose12,13, for å drive molekylær biofysikk14, og som datalagringsplattformer15.
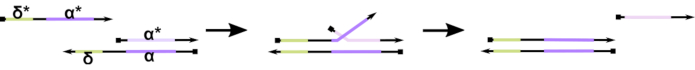
Figur 1: Mekanisme for tåholdmediert DNA-strengforskyvning. Tåholdet, δ, er en gratis, ubundet sekvens på en delvis dupleks. Når et komplementært domene (δ*) innføres på en annen tråd, fungerer det frie δ-domenet som et tåhold for hybridisering, slik at resten av tråden (ɑ*) sakte kan fortrenge konkurrenten gjennom en zipping / unzipping reversibel reaksjon kjent som strandmigrasjon. Etter hvert som lengden på δ øker, reduseres ΔG for fremoverreaksjonen, og forskyvning skjer lettere. Klikk her for å se en større versjon av denne figuren.
Til dags dato bruker flertallet av DNA-datamaskiner et veletablert motiv innen dynamisk DNA nanoteknologi kjent som tåholdmediert DNA-strandforskyvning (TMDSD, figur 1)16. Dette motivet består av en delvis dobbeltstrenget DNA (dsDNA) dupleks som viser korte “tåhold” overheng (dvs. 7- til 10 nukleotider (nt)). Nukleinsyre “input” tråder kan samhandle med de delvise dupleksene gjennom tåholdet. Dette fører til forskyvning av en av trådene fra den delvise dupleksen, og denne frigjorte strengen kan deretter tjene som inngang for nedstrøms delvise duplekser. Dermed muliggjør TMDSD signalkaskade og informasjonsbehandling. I prinsippet kan ortogonale TMDSD-motiver operere uavhengig i løsning, noe som muliggjør parallell informasjonsbehandling. Det har vært en rekke variasjoner på TMDSD-reaksjonen, for eksempel tåholdmediert DNA-strandutveksling (TMDSE)17, “lekkasjeløse” tåhold med dobbeltlange domener18, sekvens-mismatched tåhold19, og “handhold”-mediert strandforskyvning20. Disse innovative designprinsippene gir mer finjustert TMDSD-energi og dynamikk for å forbedre YTELSEN til DNA-databehandling.
Syntetiske genkretser, som transkripsjonelle genkretser, er også i stand til beregning21,22,23. Disse kretsene er regulert av proteintranskripsjonsfaktorer, som aktiverer eller undertrykker transkripsjon av et gen ved å binde seg til spesifikke regulatoriske DNA-elementer. Sammenlignet med DNA-baserte kretser har transkripsjonskretser flere fordeler. For det første har enzymatisk transkripsjon en mye høyere omsetningshastighet enn eksisterende katalytiske DNA-kretser, og genererer dermed flere kopier av utdata per enkelt kopi av inngang og gir et mer effektivt middel for signalforsterkning. I tillegg kan transkripsjonskretser produsere forskjellige funksjonelle molekyler, for eksempel aptamers eller messenger RNA (mRNA) koding for terapeutiske proteiner, som beregningsutganger, som kan utnyttes til forskjellige applikasjoner. Imidlertid er en stor begrensning av nåværende transkripsjonskretser deres mangel på skalerbarhet. Dette er fordi det er et svært begrenset sett med ortogonale proteinbaserte transkripsjonsfaktorer, og de novo-design av nye proteintranskripsjonsfaktorer forblir teknisk utfordrende og tidkrevende.
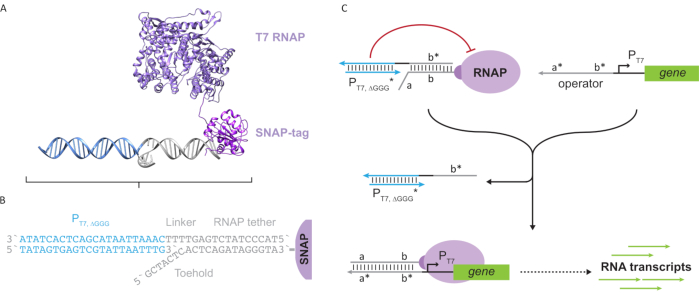
Figur 2: Abstraksjon og mekanisme for polymerasekomplekset “tether” og “cage”. (A og B) En oligonukleotidetets tether er enzymatisk merket til en T7-polymerase gjennom SNAP-tag-reaksjonen. Et bur bestående av en “faux” T7-promotor med et tether-komplementoverheng gjør det mulig å hybridisere til tether og blokkere transkripsjonsaktivitet. (C) Når operatoren (a*b*) er til stede, binder den seg til tåholdet på oligonukleotidets tether (ab) og fortrenger b* -regionen i buret, slik at transkripsjon kan forekomme. Denne figuren er endret fra Chou og Shih27. Forkortelser: RNAP = RNA polymerase. Klikk her for å se en større versjon av denne figuren.
Dette dokumentet introduserer en ny byggestein for molekylær databehandling som kombinerer funksjonaliteten til transkripsjonskretser med skalerbarheten til DNA-baserte kretser. Denne byggesteinen er en T7 RNAP som er kovalent festet med en enkeltstrenget DNA-tether (figur 2A). For å syntetisere denne DNA-tethered T7 RNAP, ble polymerasen smeltet sammen til en N-terminal SNAP-tag24 og rekombinant uttrykt i Escherichia coli. SNAP-taggen ble deretter reagert med et oligonukleotid funksjonalisert med BG-substratet. Oligonukleotid-tetheren tillater posisjonering av molekylære gjester i nærheten av polymerasen via DNA-hybridisering. En slik gjest var en konkurransedyktig transkripsjonsblokkering referert til som et “bur”, som består av en “faux” T7 promotør DNA-dupleks uten gen nedstrøms (Figur 2B). Når det bindes til RNAP via sin oligonukleotid-tether, stopper buret polymeraseaktivitet ved å utkonkurrere andre DNA-maler for RNAP-binding, noe som gjør RNAP i en “OFF” -tilstand (Figur 2C).
For å aktivere polymerasen til en “ON” tilstand, ble T7 DNA-maler med enkeltstrengede “operatør” domener oppstrøms for T7-promotoren av genet designet. Operatørdomenet (dvs. domene a*b* figur 2C) kan utformes for å fortrenge merden fra RNAP via TMDSD og plassere RNAP-proksimalt til T7-promotoren av genet, og dermed initiere transkripsjon. Alternativt ble DNA-maler også designet der operatørsekvensen var komplementær til hjelpekjernesyrestrenger som kalles “kunstige transkripsjonsfaktorer” (dvs. TFA- og TFB-tråder i figur 3A). Når begge trådene introduseres i reaksjonen, samles de på operatørstedet og oppretter et nytt pseudo-sammenhengende domene a*b*. Dette domenet kan deretter fortrenge merden via TMDSD for å starte transkripsjon (Figur 3B). Disse trådene kan leveres enten eksogent eller produseres.
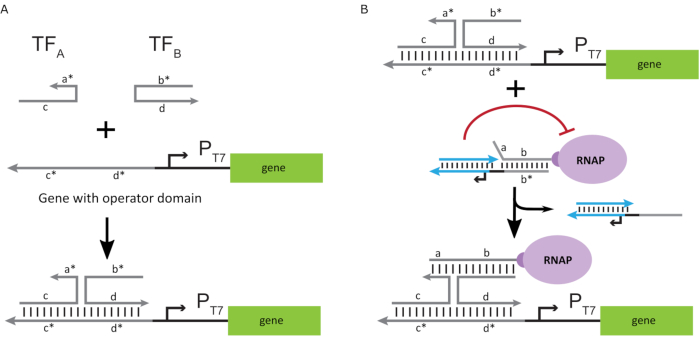
Figur 3: Selektiv programmering av polymeraseaktivitet gjennom en tre-komponent bryteraktivator. (A) Når transkripsjonsfaktorene (TFA og TFB) er til stede, binder de seg til operatørdomenet oppstrøms for promotoren, og danner en pseudo-enkeltstrenget sekvens ( a *b *) som er i stand til å fortrenge buret gjennom tåhold mediert DNA-forskyvning. (B) Dette a * b * domenet kan fortrenge buret via TMDSD for å starte transkripsjon. Denne figuren er endret fra Chou og Shih27. Forkortelser: TF = transkripsjonsfaktor; RNAP = RNA polymerase; TMDSD = tåholdmediert DNA-strengforskyvning. Klikk her for å se en større versjon av denne figuren.
Bruken av nukleinsyrebaserte transkripsjonsfaktorer for in vitro transkripsjonsregulering tillater skalerbar implementering av sofistikert kretsatferd som digital logikk, tilbakemelding og signalkaskade. For eksempel kan man bygge logikk gate kaskader ved å designe nukleinsyre sekvenser slik at transkripsjoner fra et oppstrøms gen aktiverer et nedstrøms gen. Et program som utnytter kaskade- og multipleksing gjort i stand til denne foreslåtte teknologien er utviklingen av mer sofistikerte molekylære databehandlingskretser for bærbar diagnostikk og molekylær databehandling. I tillegg kan integrering av molekylær databehandling og de novo RNA-syntesefunksjoner muliggjøre nye applikasjoner. For eksempel kan en molekylær krets utformes for å oppdage en eller en kombinasjon av brukerdefinerte RNAer som innganger og utgangsterapeutiske RNAer eller mRNAer som koder funksjonelle peptider eller proteiner for medisinske applikasjoner.
