DNA computing bruger et sæt designet oligonukleotider som medium til beregning. Disse oligonukleotider er programmeret med sekvenser til dynamisk at samle i henhold til brugerdefineret logik og reagere på specifikke nukleinsyreinput. I proof-of-concept-undersøgelser består beregningens output typisk af et sæt fluorescerende mærkede oligonuleotider, der kan påvises via gelelektroforese eller fluorescenspladelæsere. I løbet af de sidste 30 år er stadig mere komplekse DNA-beregningskredsløb blevet påvist, såsom forskellige digitale logiske kaskader, kemiske reaktionsnetværk og neurale netværk1,2,3. For at hjælpe med udarbejdelsen af disse DNA-kredsløb er matematiske modeller blevet brugt til at forudsige funktionaliteten af syntetiske genkredsløb4,5og beregningsværktøjer er udviklet til ortogonalt DNA-sekvensdesign6,7,8,9,10 . Sammenlignet med siliciumbaserede computere omfatter fordelene ved DNA-computere deres evne til at kommunikere direkte med biomolekyler, fungere i opløsning i mangel af en strømforsyning samt deres samlede kompakthed og stabilitet. Med fremkomsten af næste generations sekventering, omkostningerne ved at syntetisere DNA-computere har været faldende i de sidste to årtier med en hastighed hurtigere end Moores lov11. Anvendelser af sådanne DNA-baserede computere er nu begyndt at dukke op, såsom for sygdomsdiagnose12,13, til kraftoverførsel molekylær biofysik14,og som datalagring platforme15.
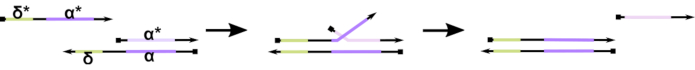
Figur 1: Mekanisme for toehold-medieret DNA-strengforskydning. Toehold, δ, er en gratis, ubundet sekvens på en delvis duplex. Når et supplerende domæne (δ*) introduceres på en anden streng, fungerer det frie δ domæne som et fodfæste for hybridisering, hvilket giver mulighed for, at resten af strengen (ɑ*) langsomt fortrænger sin konkurrent gennem en zippe / unzipping reversibel reaktion kendt som strengmigration. Efterhånden som længden af δ øges, falder ΔG for fremadrettede reaktioner, og forskydning sker lettere. Klik her for at se en større version af dette tal.
Til dato anvender de fleste DNA-computere et veletableret motiv inden for dynamisk DNA-nanoteknologi kendt som toehold-medieret DNA-strengforskydning (TMDSD, figur 1)16. Dette motiv består af en delvist dobbeltstrenget DNA (dsDNA) duplex, der viser korte “toehold” udhæng (dvs. 7- til 10 nukleotider (nt)). Nukleinsyre “input” tråde kan interagere med den delvise duplexes gennem fodfæste. Dette fører til forskydning af en af strengene fra den delvise duplex, og denne frigjorte streng kan derefter tjene som input til downstream delvise duplexes. Således muliggør TMDSD signalkascading og informationsbehandling. I princippet kan ortogonale TMDSD-motiver fungere uafhængigt i opløsning, hvilket muliggør parallel informationsbehandling. Der har været en række variationer over TMDSD-reaktionen, såsom toehold-medieret DNA-strengudveksling (TMDSE)17, “leakless” toeholds med dobbeltlange domæner18, sekvens-mismatched toeholds19og “handhold”-medieret strengforskydning20. Disse innovative designprincipper giver mulighed for mere finjusterede TMDSD-energiske og dynamikker til forbedring af DNA-databehandlingens ydeevne.
Syntetiske genkredsløb, såsom transskriptionsgenkredsløb, er også i stand til beregning21,22,23. Disse kredsløb er reguleret af protein transskription faktorer, som aktiverer eller undertrykke transskription af et gen ved at binde sig til specifikke regulatoriske DNA-elementer. Sammenlignet med DNA-baserede kredsløb har transskriptionskredsløb flere fordele. For det første har enzymatisk transskription en meget højere omsætningshastighed end eksisterende katalytiske DNA-kredsløb, hvilket genererer flere kopier af output pr. enkelt kopi af input og giver et mere effektivt middel til signalforstærkning. Derudover kan transskriptionskredsløb producere forskellige funktionelle molekyler, såsom aptamers eller messenger RNA (mRNA) kodning til terapeutiske proteiner, som beregningsoutput, som kan udnyttes til forskellige applikationer. Men en stor begrænsning af de nuværende transskriptionskredsløb er deres manglende skalerbarhed. Dette skyldes, at der er et meget begrænset sæt ortogonale proteinbaserede transskriptionsfaktorer, og de novo-design af nye proteintransskriptionsfaktorer er fortsat teknisk udfordrende og tidskrævende.
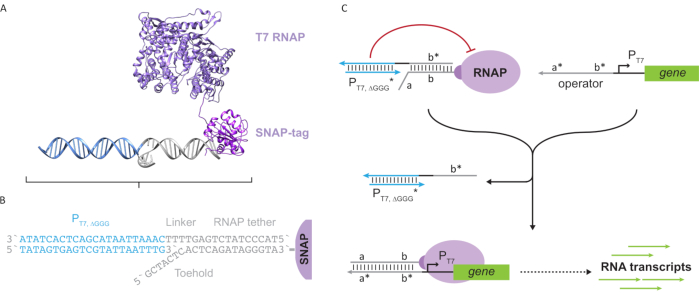
Figur 2: Abstraktion og mekanisme af “tether” og “bur” polymerase kompleks. (A og B) En oligonuleotid tether er enzymatisk mærket til en T7 polymerase gennem SNAP-tag reaktion. Et bur bestående af en “faux” T7 promotor med en tether-supplement udhæng gør det muligt at hybridisere til tøjre og blokere transskriptionelle aktivitet. (C) Når operatøren (a*b*) er til stede, binder den sig til fodfæste på oligonucleotidsbindingen(ab) og fortrænger burets b*-region, således at transskriptionen kan forekomme. Dette tal er blevet ændret fra Chou og Shih27. Forkortelser: RNAP = RNA polymerase. Klik her for at se en større version af dette tal.
Dette papir introducerer en ny byggesten til molekylær databehandling, der kombinerer funktionaliteterne i transskriptionskredsløb med skalerbarheden af DNA-baserede kredsløb. Denne byggesten er en T7 RNAP kovalent fastgjort med en enkeltstrenget DNA-tether (Figur 2A). For at syntetisere denne DNA-tøjrede T7 RNAP blev polymerase smeltet til en N-terminal SNAP-tag24 og rekombinant udtrykt i Escherichia coli. SNAP-mærket blev derefter reageret med et oligonuleotid, der blev funktionaliseret med BG-substratet. Oligonucleotid tether tillader positionering af molekylære gæster i nærheden af polymerase via DNA hybridisering. En sådan gæst var en konkurrencedygtig transskriptionsblokker kaldet et “bur”, som består af en “faux” T7 promotor DNA-duplex uden gen nedstrøms (Figur 2B). Når buret er bundet til RNAP via sinoligonucleotid tether, bremser det polymeraseaktiviteten ved at udkonkurrere andre DNA-skabeloner til RNAP-binding, hvilket gør RNAP i en “OFF”-tilstand (figur 2C).
For at aktivere polymerase til en “ON” tilstand blev T7 DNA-skabeloner med enkeltstrengede “operatør” domæner opstrøms af T7-promotoren af genet designet. Operatøren domæne (dvs. domæne a * b * Figur 2C) kan være designet til at fortrænge buret fra RNAP via TMDSD og placere RNAP proksimale til T7 promotor af genet, og dermed indlede transskription. Alternativt blev DNA-skabeloner også designet, hvor operatørsekvensen var et supplement til hjælpekernekernesalve, der kaldes “kunstige transskriptionsfaktorer” (dvs. TFA- og TFB-strenge i figur 3A). Når begge tråde introduceres i reaktionen, samles de på operatørwebstedet og opretter et nyt pseudo-sammenhængende domæne a * b *. Dette domæne kan derefter fortrænge buret via TMDSD for at indlede transskription (Figur 3B). Disse tråde kan leveres enten eksogent eller produceres.
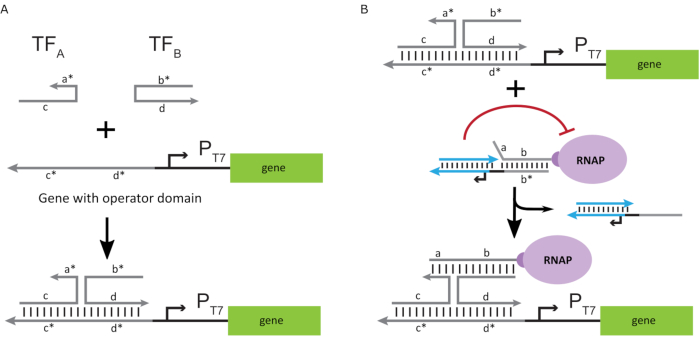
Figur 3: Selektiv programmering af polymeraseaktivitet gennem en trekomponent switch-aktivator. (A) Når transskriptionsfaktorerne (TFA og TFB) er til stede, binder de sig til operatordomænet opstrøms promotoren og danner en pseudostrenget sekvens (a*b*), der kan fortrænge buret gennem toehold medieret DNA-forskydning. (B) Dette a * b * domæne kan fortrænge buret via TMDSD at indlede transskription. Dette tal er blevet ændret fra Chou og Shih27. Forkortelser: TF = transskriptionsfaktor; RNAP = RNA polymerase; TMDSD = toehold-medieret DNA-strengforskydning. Klik her for at se en større version af dette tal.
Brugen af nukleinsyre-baserede transskription faktorer for in vitro transskriptionion regulering tillader skalerbar gennemførelse af sofistikerede kredsløb adfærd såsom digital logik, feedback, og signal cascading. For eksempel kan man bygge logik gate kaskader ved at designe nukleinsyre sekvenser således, at udskrifter fra en opstrøms gen aktivere en downstream gen. Et program, der udnytter kaskade og multiplexing gjort i stand til af denne foreslåede teknologi er udviklingen af mere sofistikerede molekylære computing kredsløb til bærbare diagnostik og molekylær databehandling. Derudover kan integration af molekylær databehandling og de novo RNA-syntesefunktioner muliggøre nye applikationer. For eksempel kan et molekylært kredsløb være designet til at detektere en eller en kombination af brugerdefinerede RNA’er som input og output terapeutiske RNA’er eller mRNAs, der kodning funktionelle peptider eller proteiner til punkt-of-care medicinske applikationer.
