Le système bipartite GAL4-UAS est le cheval de bataille de la caractérisation fonctionnelle des gènes de l’organisme modèle d’insecte Drosophila melanogaster1,2,3. Pour utiliser le système GAL4-UAS, des lignées pilotes transgéniques, exprimant le facteur de transcription de levure GAL4 sous contrôle d’une séquence régulatrice, sont croisées avec des lignes de répondeur portant un gène d’intérêt ou une construction d’interférence ARN (ARNi) contrôlée par une séquence d’activation en amont (UAS) reconnue par GAL4. La descendance de ce croisement exprime le transgène d’intérêt pour un schéma spatio-temporel dicté par le promoteur contrôlant l’expression GAL4 (Figure 1). Les phénotypes affichés par la descendance des croisements conducteur-répondeur peuvent être évalués pour élucider la fonction des gènes candidats. Bien que D. melanogaster ait été utilisé pour examiner les gènes d’autres organismes4,5,6,7, le système GAL4-UAS a maintenant été adapté pour être utilisé chez les insectes d’importance médicale et agricole afin de fournir une analyse directe des espèces d’intérêt 8,9,10,11,12,13,14.
Chez le moustique africain du paludisme, Anopheles gambiae, le système GAL4-UAS a d’abord été testé par co-transfection de lignée cellulaire9. Plusieurs constructions ont été testées pour l’efficacité dans différentes combinaisons par paires et ont constaté que 14 UAS répétés en tandem complétés par un petit intron artificiel (UAS-14i) présentaient la plus large gamme de potentiel d’activation lorsqu’ils étaient utilisés avec un panel de pilotes GAL4. Pour démontrer la fonctionnalité in vivo, ces constructions ont ensuite été utilisées pour créer deux lignées transgéniques distinctes d’An. gambiae par transformation PiggyBac8 : une ligne pilote portant GAL4 entraînée par un promoteur spécifique de l’intestin moyen, et une ligne répondeur contenant à la fois les gènes de la luciférase et de la protéine fluorescente jaune améliorée (eYFP) sous régulation des séquences UAS. L’activité de la luciférase spécifique à l’intestin et la fluorescence dans la progéniture ont indiqué que le système était efficace chez les anophèles. Depuis lors, des lignées conductrices ont été créées exprimant des transgènes dans d’autres tissus importants pour la capacité vectorielle et la résistance aux insecticides, y compris les œnocytes15 et les hémocytes16, et dans un schéma proche de l’omniprésence10. De nombreuses lignées d’UAS ont également été générées pour tester les gènes soupçonnés d’être impliqués dans le métabolisme et la résistance aux insecticides médiés par la séquestration, la synthèse des hydrocarbures cuticulaires et pour marquer par fluorescence différents types de cellules et de tissus (tableau 1). Pour les lignes répondeuses, l’intégration dirigée par site du transgène est maintenant réalisée par échange de cassettes de recombinaison catalysée ΦC31111111111 pour fixer le contexte génomique des gènes régulés par UAS. De cette façon, l’expression du transgène est normalisée en ce qui concerne l’emplacement d’insertion génomique, ce qui permet une comparaison plus précise des effets phénotypiques de différents gènes candidats.
Les lignes de répondeur créées à ce jour sont conçues pour exprimer le transgène à des niveaux élevés ou pour réduire l’expression des gènes par interférence ARN (ARNi). Habituellement, les clones d’ADNc sont fusionnés à la séquence UAS pour générer des plasmides d’expression appropriés, mais des séquences génomiques complètes sont également réalisables en supposant qu’elles ne sont pas trop grandes pour le clonage. Pour générer des constructions de silencieux, nous avons utilisé trois méthodes différentes pour obtenir des séquences inversées en tandem appropriées qui forment un ARNd en épingle à cheveux qui stimule l’ARNi. Ceux-ci ont inclus la PCR par fusion, la PCR asymétrique et la synthèse commerciale de constructions en épingle à cheveux. Chaque méthode a en commun l’inclusion d’une séquence d’introns entre les séquences inversées pour assurer la stabilité du clonage. Des plasmides répondeurs dans lesquels un gène d’intérêt/construction d’ARNi peut être inséré ont été développés15. Ces plasmides portent également les sites ΦC31 attB requis pour le RMCE (décrits dans l’article d’Adolfi accompagnant JoVE qui décrit en détail la technique RCME). Des protocoles couvrant les étapes importantes requises lors de la sélection de la séquence à insérer dans l’un de ces plasmides pour la surexpression sont inclus dans ce manuscrit. De plus, deux protocoles pour la création de construction en épingle à cheveux ARNi sont décrits et illustrés.
Lors de la création de nouvelles lignées, l’identification d’individus transgéniques rares est cruciale pour se reproduire afin d’établir et de maintenir des colonies transgéniques. Plus important encore, pour le système GAL4-UAS, il est nécessaire de distinguer les lignes d’intervention et de conducteur afin d’établir des croisements et d’identifier les descendants individuels porteurs des deux transgènes. Ceci est réalisé en utilisant différents gènes marqueurs sélectionnables dominants liés aux cassettes du conducteur et du répondeur. Le plus souvent, il s’agit de gènes marqueurs fluorescents qui peuvent être clairement distingués à l’aide de filtres optiques (par exemple, eYFP, eCFP, dsRed). Il est important que les marqueurs soient exprimés selon un schéma spatio-temporel connu et fiable, car cela facilite l’identification des anomalies et de la contamination. L’expression du gène marqueur fluorescent est régulièrement régulée par le promoteur synthétique 3xP3, qui provoque une expression spécifique des ganglions oculaires et ventraux à tous les stades du développement d’An. gambiae19. Les marqueurs fluorescents contrôlés par 3xP3 sont inclus dans tous les plasmides de transformation décrits dans cet article. Un protocole détaillant les méthodes courantes utilisées pour filtrer les lignes fluorescentes An. gambiae pupae GAL4-UAS est inclus ici.
L’un des éléments clés du système GAL4-UAS est la nécessité de traverser les lignes de conduite et d’intervention marquées différentiellement. Pour ce faire, les mâles et les femelles de chaque lignée doivent être séparés avant l’accouplement. Les adultes se distinguent facilement par la vue, cependant, pour établir des croisements génétiques, il est judicieux de séparer les sexes avant l’émergence de l’adulte pour s’assurer que l’accouplement n’a pas eu lieu. La différence de taille générale entre les pupes an. gambiae mâles et femelles est trop variable pour être une méthode efficace et fiable de détermination du sexe20. Au lieu de cela, des différences morphologiques claires dans les organes génitaux externes fournissent une base fiable pour le sexage chez An. gambiae. Dans cet article, nous décrivons une méthode fiable pour sexer les pupes d’An. gambiae afin de mettre en place des croisements appropriés.
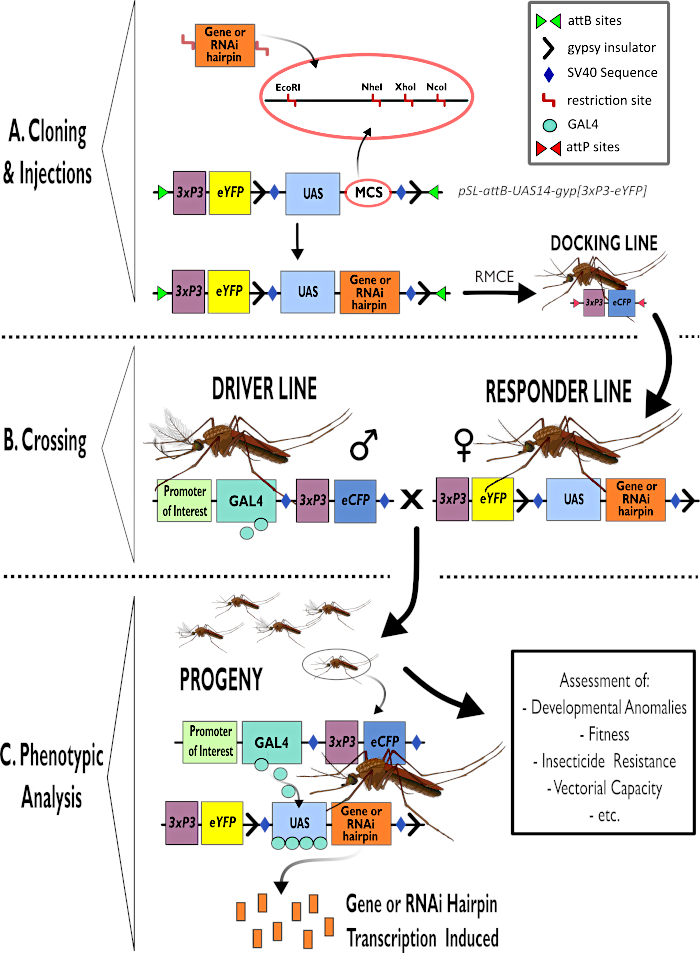
Figure 1 – Représentation schématique du processus d’utilisation du système bipartite GAL4-UAS dans Anopheles gambiae. (A) Les principaux composants d’un exemple de vecteur (pSL-attB-UAS14-gyp[3xp3-eYFP]) sont représentés, détaillant les sites de restriction disponibles (EcoRI, NheI, XhoI et NcoI) dans les multiples sites de clonage qui peuvent être utilisés pour insérer la construction en épingle à cheveux ou la séquence codante pour le gène d’intérêt. La structure de la ligne d’amarrage est également représentée. (B) L’étape de croisement est illustrée en indiquant l’utilisation d’hommes de la ligne de conduite (transportant le conducteur GAL4 par un promoteur d’intérêt et eCFP piloté par le promoteur 3xP3) et de femelles de la ligne de réponse (portant le gène d’intérêt ou la construction en épingle à cheveux contrôlée par un promoteur UAS et un marqueur eYFP contrôlé par le promoteur 3xP3). (C) Une représentation schématique de GAL4 conduisant l’expression du gène d’intérêt dans la descendance du croisement dans B et une liste de certains des phénotypes typiques qui sont évalués. Abréviations : Site de clonage multiple (MCS), échange de cassettes médié par la recombinase (RMCE), séquence d’activateur en amont (UAS), protéine fluorescente jaune améliorée (eYFP), protéine fluorescente cyan améliorée (eCFP). Veuillez cliquer ici pour voir une version agrandie de cette figure.
C’est l’utilisation de croisements qui fournit la nature bipartite du système GAL4-UAS, qui présente des avantages distincts par rapport aux approches plus linéaires. Par exemple, beaucoup plus de combinaisons de lignées conductrices et répondeuses peuvent être évaluées que ce qui serait faisable si une nouvelle lignée transgénique devait être générée et maintenue pour chaque combinaison promoteur/gène. Plus important encore, il permet l’analyse de gènes qui produisent des phénotypes mortels ou stériles lorsque leur expression est perturbée et qui sont difficiles à créer / maintenir dans un système linéaire. De tels phénotypes mortels peuvent se manifester à tous les stades de développement, en fonction de la fonction du gène et de l’expression spatio-temporelle, mais sont le plus souvent observés au cours du développement embryonnaire. La visualisation du développement embryonnaire de moustique nécessite l’élimination du chorion opaque qui recouvre les œufs. En suivant les méthodes décrites dans Trpiš (1970)21 et Kaiser et al. (2014)22, nous décrivons les protocoles que nous utilisons pour fixer les embryons, tout en maintenant l’intégrité structurelle, et le blanchiment pour éliminer l’endochorion qui permet la visualisation et l’imagerie microscopiques.
