O sistema bipartite GAL4-UAS é o cavalo de trabalho da caracterização funcional de genes no organismo modelo de insetos Drosophila melanogaster1,2,3. Para usar o sistema GAL4-UAS, as linhas de driver transgênicas, expressando o fator de transcrição da levedura GAL4 sob controle de uma sequência regulatória, são cruzadas com linhas de resposta carregando um gene de interesse ou interferência de RNA (RNAi) construída controlada por uma Sequência de Ativação Upstream (UAS) reconhecida pela GAL4. A descendência dessa cruz expressa o transgene de interesse em um padrão espítetempo ditado pelo promotor que controla a expressão GAL4 (Figura 1). Fenótipos exibidos por descendentes de cruzes de respondente condutor podem ser avaliados para elucidar a função dos genes candidatos. Embora d. melanogaster tenha sido usado para examinar genes de outros organismos4,5,6,7, o sistema GAL4-UAS foi agora adaptado para uso em insetos de importância médica e agrícola para fornecer análise direta nas espécies de interesse 8,9,10,11,12,13,14.
No mosquito africano da malária, Anopheles gambiae, o sistema GAL4-UAS foi testado pela primeira vez pela co-transfecção da linha celular9. Construções múltiplas foram avaliadas para eficiência em diferentes combinações de pairwise e descobriram que 14 UAS repetidas com um pequeno intron artificial (UAS-14i) exibiam a maior gama de potencial de ativação quando usado com um painel de drivers GAL4. Para demonstrar a funcionalidade in vivo, essas construções foram então usadas para criar duas linhas transgênicas de An. gambiae separadas pela Transformação PiggyBac8: uma linha de driver que transporta GAL4 conduzida por um promotor específico midgut, e uma linha de resposta contendo tanto a luciferase quanto os genes de proteína fluorescente amarela aprimorada (eYFP) sob regulação das sequências UAS. A atividade de luciferase específica intestinal e a fluorescência na prole indicaram que o sistema era eficiente em Anopheles. Desde então, foram criadas linhas de condutores expressando transgenes em outros tecidos importantes para a capacidade vetorial e resistência a inseticidas, incluindo oenocitos15 e hemócitos16, e em um padrão próximo ao onipresente10. Numerosas linhas de UAS também foram geradas para avaliar genes considerados envolvidos no metabolismo e sequestração mediada resistência a inseticidas, síntese de hidrocarbonetos cuticulares e para marcar fluorescente diferentes tipos de células e tecidos (Tabela 1). Para as linhas de resposta, a integração direcionada ao local do transgene é agora realizada pelo ΦC31 catalisado troca de cassete17,18 para corrigir o contexto genômico dos genes regulados pela UAS. Dessa forma, a expressão transgênica é normalizada em relação à localização de inserção genômica, permitindo uma comparação mais precisa dos efeitos fenotípicos de diferentes genes candidatos.
As linhas de respondente criadas até o momento são projetadas para expressar o transgene em níveis elevados ou reduzir a expressão genética através da interferência de RNA (RNAi). Normalmente, os clones cDNA são fundidos à sequência UAS para gerar plasmídeos de expressão adequados, porém sequências genômicas completas também são viáveis assumindo que eles não são muito grandes para clonagem. Para gerar construções de silenciamento, utilizamos três métodos diferentes para obter sequências invertidas tandem adequadas que formam o dsRNA que estimula o RNAi. Estes incluíram PCR de fusão, PCR assimétrico e síntese comercial de construções de grampos de cabelo. Comum a cada método é a inclusão de uma sequência intron entre as sequências invertidas para fornecer estabilidade de clonagem. Plasmids de resposta nos quais um gene de interesse/construção RNAi pode ser inserido foram desenvolvidos15. Esses plasmídeos também carregam os locais de attB ΦC31 necessários para rmce (descrito em Adolfi acompanhando o artigo jove que descreve a técnica RCME em detalhes). Protocolos que abrangem as etapas importantes necessárias ao selecionar a sequência para inserção em um desses plasmídeos para superexpressão estão incluídos neste manuscrito. Além disso, dois protocolos para criação de construção de grampos rnai são descritos e ilustrados.
Ao criar novas linhas, a identificação de indivíduos transgênicos raros é crucial para procriar para estabelecer e manter colônias transgênicas. Mais importante para o sistema GAL4-UAS, é necessário distinguir as linhas de respondente e motorista para estabelecer cruzes e identificar descendentes individuais que carregam ambos transgenes. Isso é conseguido usando diferentes genes de marcadores selecionáveis dominantes ligados às fitas do motorista e do respondente. Mais comumente estes são genes marcadores fluorescentes que são claramente distinguíveis usando filtros ópticos (por exemplo, eYFP, eCFP, dsRed). É importante que os marcadores sejam expressos em um padrão espíte conhecido e confiável, pois isso facilita a identificação de anormalidades e contaminação. A expressão genética do marcador fluorescente é rotineiramente regulada pelo promotor 3xP3 sintético, que causa expressão específica de gânglios oculares e ventral em todas as etapas do desenvolvimento de An. gambiae19. Marcadores fluorescentes controlados por 3xP3 estão incluídos em todos os plasmídeos de transformação descritos neste artigo. Um protocolo detalhando os métodos comuns usados para tela fluorescente Uma. gambiae pupae GAL4-UAS está incluído aqui.
Um dos elementos-chave do sistema GAL4-UAS é a necessidade de cruzar as linhas de driver e responder marcadas diferencialmente. Para fazer este macho e fêmeas de cada linha deve ser separado antes do acasalamento. Os adultos são facilmente distinguíveis pela visão, no entanto, para estabelecer cruzes genéticas é sensato separar os sexos antes do surgimento de adultos para garantir que o acasalamento não tenha ocorrido. A diferença geral de tamanho entre macho e fêmea An. gambiae pupae é muito variável para ser um método eficiente e confiável de determinação sexual20. Em vez disso, diferenças morfológicas claras na genitália externa fornecem uma base confiável para o sexing em An. gambiae. Neste artigo, descrevemos um método confiável para sexar An. gambiae pupae para configurar cruzes apropriadas.
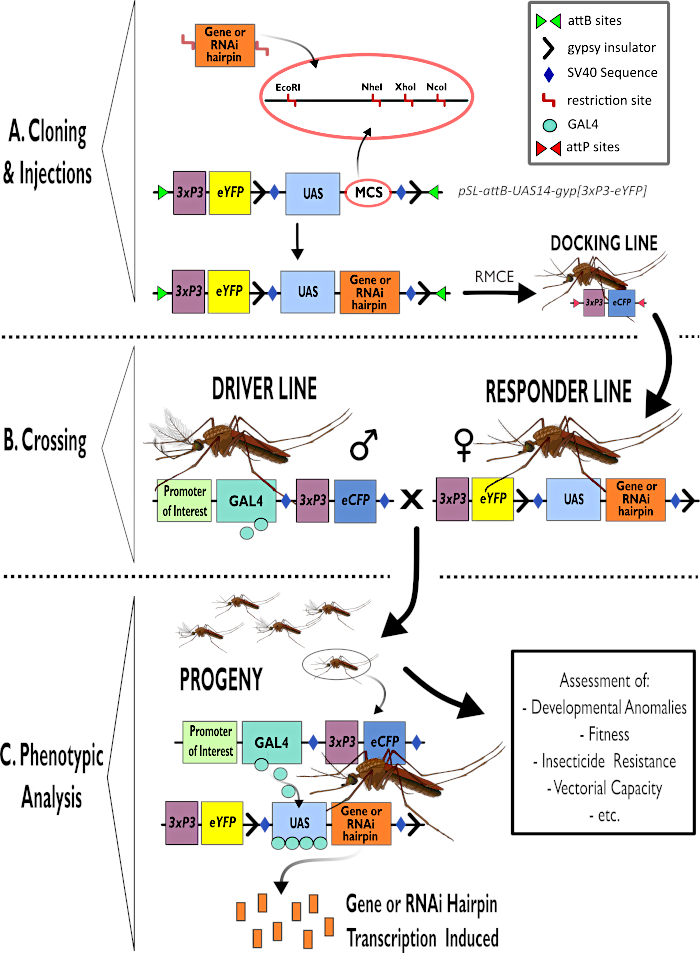
Figura 1 – Representação diagramática do processo para o uso do sistema bipartite GAL4-UAS em Anopheles gambiae. (A) Os principais componentes de um vetor de exemplo (pSL-attB-UAS14-gyp[3xp3-eYFP]) são retratados, detalhando os locais de restrição disponíveis (EcoRI, NheI, XhoI e NcoI) dentro dos múltiplos locais de clonagem que são adequados para uso para inserir a construção de grampos ou sequência de codificação para o interesse genético. A estrutura da linha de acoplamento também é retratada. (B) A etapa de travessia é ilustrada indicando o uso de machos da linha de motorista (transportando condutor gal4 por um promotor de interesse e eCFP conduzido pelo promotor 3xP3) e fêmeas da linha de resposta (carregando o gene de interesse ou construção de grampos controlado por um promotor da UAS e um marcador eYFP controlado pelo promotor 3xP3). (C) Uma representação diagramática da expressão de condução GAL4 do gene de interesse na prole da cruz em B e uma lista de alguns dos fenótipos típicos que são avaliados. Abreviaturas: Multiple Cloning Site (MCS), Recombinase mediated cassette exchange (RMCE), Upstream Activator Sequence (UAS), proteína fluorescente amarela aprimorada (eYFP), proteína fluorescente ciano aprimorada (eCFP). Clique aqui para ver uma versão maior desta figura.
É o uso de cruzes que fornece a natureza bipartite do sistema GAL4-UAS, que tem vantagens distintas sobre abordagens mais lineares. Por exemplo, muitas mais combinações de linhas de driver e respondente podem ser avaliadas do que seria viável se uma nova linha transgênica tivesse que ser gerada e mantida para cada combinação promotor/gene. Mais importante, permite a análise de genes que produzem fenótipos letais ou estéreis quando sua expressão é perturbada, que são difíceis de criar/manter em um sistema linear. Tais fenótipos letais podem se manifestar em todos os estágios de desenvolvimento, dependendo da função genética e expressão espacial, mas são mais frequentemente observados durante o desenvolvimento embrionário. Visualizar o desenvolvimento de embriões de mosquitos requer a limpeza da chorão opaca que reveste os ovos. Seguindo os métodos descritos em Trpiš (1970)21 e Kaiser et al. (2014)22, descrevemos os protocolos que usamos para corrigir embriões, mantendo a integridade estrutural e o branqueamento para limpar a endochorion que permite visualização e imagem microscópicas.
