Бактерии используют несколько небольших сигнальных молекул для адаптации к постоянно меняющимся средам1,2. Например, аутоиндукруторы, N-ацилгомосериновые лактоны и их модифицированные олигопептиды, опосредовывают межклеточную связь между бактериями для координации поведения популяции, явление, известное как ощущение кворума2. Другой группой малых сигнальных молекул является NSM, включая широко изученный циклический аденозинмонофосфат (цАМФ), циклический ди-АМФ, циклический ди-гуанозинмонофосфат (циклический ди-GMP) и гуанозин пента- и тетрафосфаты (p)ppGpp1. Бактерии производят эти NSM в ответ на множество различных стрессовых состояний. После производства эти молекулы связываются со своими белками-мишенями и регулируют несколько различных физиологических и метаболических путей, чтобы справиться с возникающими стрессами и повысить выживаемость бактерий. Поэтому идентификация белков-мишеней является неизбежной предпосылкой для расшифровки молекулярных функций этих малых молекул.
Последнее десятилетие стало свидетелем бума знаний об этих небольших сигнальных молекулах, в основном благодаря нескольким техническим инновациям, которые раскрыли целевые белки этих малых молекул. К ним относятся метод захвата соединения3,4,5и дифференциальное радиальное капиллярное действие лигандного анализа (DRaCALA)6, которые будут обсуждаться в этой статье.
Изобретенный Винсентом Ли и его коллегами в 2011году 6,DRaCALA использует способность нитроцеллюлозной мембраны дифференциально секвестрировать свободные и связанные с белком лиганды. Молекулы, такие как белки, не могут диффундировать на нитроцеллюлозной мембране, в то время как небольшие лиганды, такие как NSM, способны. Смешивая NSM(например,ppGpp) с тестируемым белком и обличивая их на мембране, можно ожидать двух сценариев(рисунок 1):Если (p)ppGpp связывается с белком, радиомаркированный (p)ppGpp будет удерживаться в центре пятна белком и не будет диффундировать наружу, давая интенсивную маленькую точку(т.е., сильный радиоактивный сигнал) под фосфоритатором. Однако, если (p)ppGpp не связывается с белком, он будет свободно диффундировать наружу, производя большое пятно с однородным фоновым радиоактивным сигналом.
Кроме того, DRaCALA может обнаружить взаимодействие между небольшой молекулой и неочищенным белком в цельном клеточном лизате, если белок присутствует в достаточном количестве. Эта простота позволяет использовать DRaCALA для быстрой идентификации белковых мишеней с помощью библиотеки экспрессии ORFeome. Действительно, целевые белки цАМФ7,циклический ди-АМФ8,циклический ди-GMP9,10и (p)ppGpp11,12,13 были систематически идентифицированы с помощью DRaCALA. В этой видеостатье используется (p)ppGpp в качестве примера для демонстрации и описания критических шагов и соображений при успешном проведении скрининга DRaCALA. Следует отметить, что более подробное описание DRaCALA14 настоятельно рекомендуется прочитать в сочетании с этой статьей перед выполнением DRaCALA.
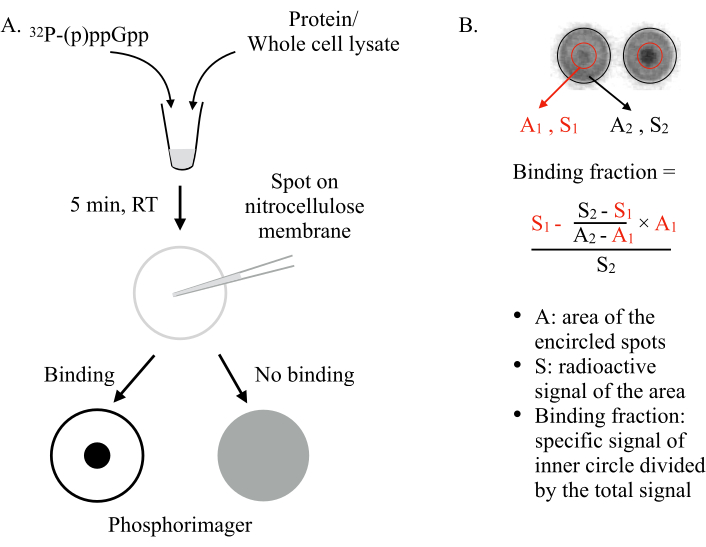
Рисунок 1:Принцип DRaCALA. (A) Схема анализа DRaCALA. Подробности см. в тексте. (B)Количественная оценка и расчет фракции связывания. Подробности см. в тексте. Вкратце, пятна DRaCALA будут проанализированы путем рисования двух кругов, которые ограничивают все пятно и внутреннюю темную точку(т. Е.Сохраненный (p)ppGpp из-за связывания тестируемого белка). Специфическим связывающим сигналом является радиоактивный сигнал внутреннего круга (S1) после вычитания неспецифического фонового сигнала (вычисляется по A1 × ((S2-S1)/(A2-A1))). Фракция связывания представляет специфический сигнал связывания, деленный на общий радиоактивный сигнал (S2). Сокращения: DRaCALA = дифференциальное радиальное капиллярное действие лигандного анализа; (p)ppGpp = гуанозин пента- и тетрафосфаты; RT = комнатная температура. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Работа поддерживается грантом проекта NNF (NNF19OC0058331) для YEZ и исследовательской и инновационной программой Европейского Союза Horizon 2020 в рамках грантового соглашения Марии Склодовской-Кюри (Nº 801199) для MLS.

