La comprensión de las membranas celulares y su función ha evolucionado notablemente en las últimas décadas. La visión anterior de las membranas celulares como bicapas lipídicas pasivas que definen los límites celulares y albergan proteínas de membrana1 se ha transformado gradualmente en un modelo dinámico en el que las bicapas lipídicas desempeñan un papel importante en la regulación de los procesos biológicos vitales, incluida la señalización celular, el intercambio molecular y la función de las proteínas, por nombrar algunos2,3,4,5,6. Esta comprensión de que las membranas celulares son altamente dinámicas, en constante remodelación y redistribución molecular, ha impulsado exploraciones científicas más allá de las estructuras de equilibrio de las membranas7,8,9. En consecuencia, se han desarrollado múltiples enfoques para estudiar los diversos modos dinámicos en las membranas lipídicas biológicas y bioinspiradas. Hasta la fecha, la mayoría de estos estudios se han centrado principalmente en los movimientos moleculares difusivos10,11,12,13 y las fluctuaciones macroscópicas de la forma14,15,16,dejando un vacío significativo en la comprensión de la dinámica intermedia de la membrana, es decir, las fluctuaciones colectivas de los conjuntos de lípidos que consisten en unos pocos 10-100 de moléculas de lípidos. Estas dinámicas ocurren en escalas de longitud de pocas decenas a pocas 100 Å y en escalas de tiempo de sub-ns a unos pocos cientos de ns (ver Figura 1),referidas aquí como escalas mesoscópicas. De hecho, es en estas escalas que la actividad biológica clave tiene lugar a nivel de membrana17. Esto incluye la brotación viral18,la canalización19y las interacciones membrana-proteína20. También es importante señalar que el panorama energético de las proteínas de membrana21,22 muestra que los cambios conformacionales en las proteínas, necesarios para su papel regulador, ocurren en las escalas de tiempons 23 de fluctuaciones colectivas de la membrana, enfatizando aún más la importancia de la dinámica mesoscópica en la función biológica de las membranas celulares y sus análogos bioinspirados20. Este artículo se centra en los dos modos dinámicos mesoscópicos primarios en las membranas lipídicas, a saber, las fluctuaciones de flexión y las fluctuaciones de espesor.
El principal desafío al sondear directamente estos modos de fluctuación es la dificultad de acceder simultáneamente a sus escalas espaciales y temporales utilizando métodos de espectroscopia estándar. El otro desafío es que las técnicas de contacto directo podrían afectar las mismas fluctuaciones que se supone que miden16. Esto se ve agravado por la complejidad composicional y estructural de las membranas biológicas24,25,lo que resulta en características de membrana no homogéneas, incluida la formación de dominios lipídicos26,27,28,29,30 y la asimetría de membrana31,32,33, que exigen sondas selectivas para comprender la dinámica de diferentes características de la membrana. Afortunadamente, estos desafíos pueden superarse con métodos de espectroscopia de neutrones no invasivos, como el eco de espín de neutrones (NSE), que inherentemente accede a las escalas de longitud y tiempo requeridas, y permiten además estudios de características selectivas de la membrana sin cambiar su entorno fisicoquímico34. De hecho, en los últimos años la espectroscopia NSE se ha convertido en una sonda única y poderosa de dinámica colectiva demembranas 35. Los resultados de los estudios de NSE sobre membranas lipídicas han producido nuevos conocimientos sobre las propiedades mecánicas36,37 y viscoelásticas38,39 de las membranas lipídicas y han arrojado nueva luz sobre su papel potencial en la función biológica40,41.
La técnica de espectroscopia NSE se basa en un diseño de instrumento interferométrico, propuesto por primera vez por Mezei42,utilizando una serie de aletas de espín y bobinas magnéticas para controlar la precesión del espín de neutrones a medida que los neutrones atraviesan el instrumento. El diseño se basa en el reflejo magnético de los elementos del campo magnético con respecto a la posición de la muestra (Figura 1A). Esto implica que en ausencia de intercambio de energía entre el neutrón y la muestra, el neutrón realiza el mismo número de precesiones de espín, en direcciones opuestas, en la primera y segunda mitad del instrumento (nótese el π-flipper entre las dos bobinas de precesión). Como resultado, el estado de espín final del neutrón permanece sin cambios en relación con el estado inicial, un fenómeno conocido como espín-eco (ver neutrón transparente en la Figura 1A). Sin embargo, cuando el neutrón interactúa energéticamente con la muestra, el intercambio de energía modifica el número de precesiones de espín en la segunda mitad del instrumento, lo que lleva a un estado de espín final diferente (ver Figura 1A). Esto se detecta experimentalmente como una pérdida en la polarización, como se mostrará más adelante en este artículo. Para obtener más detalles sobre la técnica NSE, se remite al lector a los documentos técnicos dedicados42,43,44,45.
Aquí, se presenta una descripción simplificada para proporcionar una estimación aproximada de la longitud y las escalas de tiempo accesibles con NSE. Las escalas de longitud están determinadas por el rango de transferencias de vector de onda alcanzables, Q = 4π sin θ/λ, donde 2θ es el ángulo de dispersión y λ es la longitud de onda del neutrón. Se puede ver que Q está establecido por el rango de longitud de onda y la extensión de rotación del segundo brazo del espectrómetro (ver Figura 1A). Un rango Qtípico en los espectrómetros NSE es ~ 0.02-2 Å-146,47y hasta 0.01-4 Å-1 con actualizaciones recientes48,49, correspondientes a escalas espaciales de ~ 1-600 Å. Por otro lado, la escala de tiempo accesible se calcula a partir del ángulo de precesión total (o fase) adquirido por el neutrón dentro de las bobinas de precesión magnética, y se encuentra que esde 50: 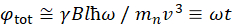 . En esta expresión, t es el tiempo de Fourier definido como
. En esta expresión, t es el tiempo de Fourier definido como 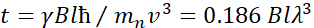 , donde es la relación
, donde es la relación 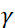 giromagnética de neutrones,
giromagnética de neutrones, 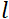 es la longitud de la bobina y es la fuerza del campo magnético de
es la longitud de la bobina y es la fuerza del campo magnético de 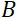 la bobina. Vale la pena señalar que el tiempo de Fourier es una cantidad que depende estrictamente de la geometría del instrumento, la intensidad del campo magnético y la longitud de onda de los neutrones. Por ejemplo, utilizando neutrones de longitud de onda
la bobina. Vale la pena señalar que el tiempo de Fourier es una cantidad que depende estrictamente de la geometría del instrumento, la intensidad del campo magnético y la longitud de onda de los neutrones. Por ejemplo, utilizando neutrones de longitud de onda 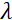 = 8 Å y ajustes del instrumento de = 1,2 m y = 0,4 T, el tiempo de Fourier se calcula en t ~ 50 ns. Experimentalmente, el tiempo de Fourier se ajusta cambiando la corriente en las bobinas de precesión (es decir, la intensidad del campo magnético) o utilizando diferentes longitudes de onda de neutrones, lo que resulta en escalas de tiempo típicas de NSE de ~ 1 ps a 100 ns. Sin embargo, las actualizaciones recientes en los espectrómetros NSE han permitido el acceso a tiempos Fourier más largos, hasta ~ 400 ns en el espectrómetro J-NSE-Phoenix en el Heinz Maier-Leibnitz Zentrum51 y el espectrómetro SNS-NSE en el Laboratorio Nacional Oak Ridge48,y hasta ~ 1,000 ns en el espectrómetro IN15 NSE en el Institut Laue-Langevin (ILL)49.
= 8 Å y ajustes del instrumento de = 1,2 m y = 0,4 T, el tiempo de Fourier se calcula en t ~ 50 ns. Experimentalmente, el tiempo de Fourier se ajusta cambiando la corriente en las bobinas de precesión (es decir, la intensidad del campo magnético) o utilizando diferentes longitudes de onda de neutrones, lo que resulta en escalas de tiempo típicas de NSE de ~ 1 ps a 100 ns. Sin embargo, las actualizaciones recientes en los espectrómetros NSE han permitido el acceso a tiempos Fourier más largos, hasta ~ 400 ns en el espectrómetro J-NSE-Phoenix en el Heinz Maier-Leibnitz Zentrum51 y el espectrómetro SNS-NSE en el Laboratorio Nacional Oak Ridge48,y hasta ~ 1,000 ns en el espectrómetro IN15 NSE en el Institut Laue-Langevin (ILL)49.
Además del acceso directo a la escala de tiempo y longitud de la dinámica de membranas, NSE tiene las capacidades inherentes de la sensibilidad a los isótopos de neutrones52. Específicamente, la capacidad de los neutrones para interactuar de manera diferente con los isótopos del hidrógeno, el elemento más abundante en los sistemas biológicos, da como resultado una densidad de longitud de dispersión de neutrones diferente,34 o NSLD (el equivalente al índice óptico de refracción50),cuando el protio es sustituido por deuterio. Esto permite un enfoque conocido como variación de contraste, que se usa comúnmente para resaltar características específicas de la membrana u ocultar otras; este último escenario se conoce como coincidencia de contraste. Una aplicación frecuente de variación/coincidencia de contraste es la sustitución de agua (NSLD = -0.56 × 10-6 Å-2) por agua pesada o D2O (NSLD = 6.4 × 10-6 Å -2) para amplificar la señal de neutrones de las membranas lipídicas protiatadas (NSLD ~ 0 × 10-6 Å-2). Este enfoque es altamente efectivo en estudios de estructura de membrana porque la penetración de D2O en la región del grupo de cabeza de la membrana permite la determinación precisa de los espesores de la membrana (ver Figura 2A,panel izquierdo) y de la ubicación de diferentes subgrupos lipídicos cuando se aplican modelos más sofisticados53,54. Este artículo destaca algunos ejemplos sobre el uso de la variación de contraste para estudios de dinámica colectiva en membranas biomiméticas y características de membrana seleccionadas.
Aquí, la efectividad de NSE para proporcionar información única sobre las propiedades dinámicas y funcionales de la membrana se ilustra a través de ejemplos tangibles de estudios de NSE en sistemas de membrana lipídica modelo y biológicamente relevantes con énfasis en la dinámica de mesoescala en membranas independientes, en forma de suspensiones liposomales. Para las mediciones de NSE de la dinámica de la membrana en el plano, se remite al lector a publicaciones dedicadas a la espectroscopia de espín-eco de neutrones de incidencia de pastoreo (GINSES)55,56 y otros estudios de pilas de membrana multilamelar alineadas57,58,59,60.
Para simplificar, este artículo destaca tres esquemas diferentes de deuteración de membrana ilustrados en un sistema de bicapa lipídica de formación de dominios o separación de fases bien estudiado de mezclas de 1,2-ditiristoil-sn-glicero-3-fosfocolina (DMPC) y 1,2-distearoil-sn-glicero-3-fosfocolina (DSPC)61,62. Los dos lípidos se caracterizan por un desajuste en la longitud de su cadena de hidrocarburos (14 carbonos/cola en DMPC vs 18 carbonos/cola en DSPC) y su temperatura de transición gel-fluido (Tm, DMPC = 23 °C vs Tm, DSPC = 55 °C). Esto da como resultado la separación de fases laterales en membranas DMPC:DSPC a temperaturas entre las temperaturas de transición superior e inferior de la mezcla63. Los esquemas de deuteración considerados aquí se eligen para demostrar los diferentes modos dinámicos accesibles en las mediciones de NSE en membranas liposomales, a saber, fluctuaciones de flexión, fluctuaciones de espesor y fluctuaciones selectivas de flexión / espesor de dominios laterales. Todas las composiciones lipídicas se informan para las bicapas DMPC:DSPC preparadas a una fracción molar de 70:30, utilizando variantes protiated y perdeuteradas disponibles comercialmente de DMPC y DSPC. Todos los pasos de preparación de la muestra se basan en 4 mL de suspensión liposomal, en D2O, con una concentración de lípidos de 50 mg/mL, para una masa lipídica total de Mtot = 200 mg por muestra.
