تطور فهم أغشية الخلايا ووظيفتها بشكل ملحوظ على مدى العقود القليلة الماضية. وقد تحولت النظرة السابقة لأغشية الخلايا كثنائيات الدهون السلبية التي تحدد حدود الخلايا وبروتينات غشاء المنزل1 تدريجيا إلى نموذج ديناميكي تلعب فيه الطبقات ثنائية الدهون دورا مهما في تنظيم العمليات البيولوجية الحيوية ، بما في ذلك الإشارات الخلوية والتبادل الجزيئي ووظيفة البروتين – على سبيل المثاللا الحصر2و3و4و5و6. هذا الإدراك بأن أغشية الخلايا ديناميكية للغاية ، وتخضع باستمرار لإعادة العرض وإعادة التوزيع الجزيئي ، وقد حث الاستكشافات العلمية وراء هياكل التوازن للأغشية7و8و9. وبناء على ذلك، تم تطوير نهج متعددة لدراسة مختلف الأنماط الديناميكية في الأغشية الدهنية البيولوجية والحيوى. حتى الآن، ركزت غالبية هذه الدراسات في المقام الأول على نشر الحركات الجزيئية10،11،12،13 وتقلبات الشكل العياني14،15،16، مما يترك فجوة كبيرة في فهم ديناميات الأغشية المتوسطة ، أي التقلبات الجماعية لجمعيات الدهون التي تتكون من عدد قليل من 10-100s من جزيئات الدهون. تحدث هذه الديناميكيات على مقاييس طول من عشرات قليلة إلى عدد قليل من 100 Å وعلى مدى المقاييس الزمنية من ns الفرعية إلى بضع مئات من ns (انظر الشكل 1)، ويشار إليها هنا باسم جداول التنظير. هو حقا على هذا ميزان أن نشاط رئيسية أحيائية يتم في الغشاء مستوى17. وهذا يشمل الفيروسية الناشئة18، قناة gating19، والتفاعلات الغشاء البروتين20. من المهم أيضا الإشارة إلى أن مشهد الطاقة للبروتينات الغشائية21،22 يظهر أن التغيرات التوافقية في البروتينات – الضرورية لدورها التنظيمي – تحدث على مدى المقاييس الزمنية23 من تقلبات الأغشية الجماعية ، مما يؤكد كذلك على أهمية الديناميكيات التنظيرية في الوظيفة البيولوجية لأغشية الخلايا ونظائرها ذات الأغشية الحيوية20. تركز هذه الورقة على الوضعين الديناميكيين الأساسيين بالمنظار في الأغشية الدهنية ، وهما تقلبات الانحناء وتقلبات السماكة.
ويتمثل التحدي الرئيسي في التحقيق المباشر في أوضاع التقلبات هذه في صعوبة الوصول في الوقت نفسه إلى مقاييسها المكانية والزمنية باستخدام أساليب التحليل الطيفي القياسية. والتحدي الآخر هو أن تقنيات الاتصال المباشر يمكن أن تؤثر على نفس التقلبات التي يقصد بها قياس16. ومما يزيد من تفاقم هذا التعقيد التركيبي والهيكلي للأغشية البيولوجية24،25، مما يؤدي إلى ميزات غشاء غير متجانسة ، بما في ذلك تشكيل المجال الدهني26،27،28،29،30 وعدم تناسق الأغشية31،32،33– مما يتطلب تحقيقات انتقائية لفهم ديناميات ميزات الأغشية المختلفة. لحسن الحظ ، يمكن التغلب على هذه التحديات باستخدام طرق التحليل الطيفي النيوتروني غير الغازية ، مثل صدى الدوران النيوتروني (NSE) ، والتي تدخل بطبيعتها إلى الطول والمقاييس الزمنية المطلوبة ، وتمكن من إجراء دراسات حول ميزات الغشاء الانتقائي دون تغيير بيئتها الفيزيائيةالكيميائية 34. في الواقع ، على مدى السنوات القليلة الماضية تطور التحليل الطيفي NSE إلى مسبار فريد وقوي من ديناميكيات الغشاء الجماعي35. وقد أنتجت نتائج دراسات NSE على الأغشية الدهنية رؤى جديدة فيالميكانيكية 36،37 وviscoelastic38،39 خصائص الأغشية الدهنية وألقت ضوءا جديدا على دورها المحتمل في الوظيفة البيولوجية40،41.
تعتمد تقنية التحليل الطيفي NSE على تصميم أداة التداخل ، الذي اقترحه Mezei42لأول مرة ، باستخدام سلسلة من الزعانف الدوارة واللفائف المغناطيسية للتحكم في مقدمة الدوران النيوتروني أثناء اجتياز النيوترونات للجهاز. يعتمد التصميم على النسخ المتطابق المغناطيسي لعناصر المجال المغناطيسي فيما يتعلق بموضع العينة(الشكل 1A). وهذا يعني أنه في غياب تبادل الطاقة بين النيوترون والعينة ، يؤدي النيوترون نفس العدد من السبق الدوراني ، في اتجاهين متعاكسين ، في النصف الأول والثاني من الجهاز (لاحظ π الزعانف بين لفائف السبق). ونتيجة لذلك، تبقى حالة الدوران النهائية للنيوترون دون تغيير بالنسبة للدولة الأولية – وهي ظاهرة يشار إليها باسم الدوران صدى (انظر النيوترون الشفاف في الشكل 1A). ومع ذلك ، عندما يتفاعل النيوترون بنشاط مع العينة ، يعدل تبادل الطاقة عدد السبقات الدورانية في النصف الثاني من الجهاز ، مما يؤدي إلى حالة دوران نهائية مختلفة (انظر الشكل 1A). يتم الكشف عن هذا تجريبيا كخسارة في الاستقطاب ، كما سيظهر لاحقا في هذه الورقة. لمزيد من التفاصيل حول تقنية NSE ، يشار إلى القارئ إلى الأوراق الفنيةالمخصصة 42،43،44،45.
هنا، يتم تقديم وصف مبسط لتقديم تقدير تقريبي للطول والجداول الزمنية التي يمكن الوصول إليها مع NSE. يتم تحديد موازين الطول من خلال نطاق عمليات نقل الموجات القابلة للتحقيق، Q = 4π sin ο/λ، حيث 2ο هي زاوية التشتت و λ هو الطول الموجي النيوتروني. يمكن للمرء أن يرى أن يتم تعيين Q من قبل نطاق الطول الموجي ومدى دوران الذراع الثاني للمطياف (انظر الشكل 1A). نموذجي Q-rangeعلى مطياف NSE هو ~0.02-2 Å-146,47, وحتى 0.01-4 Å-1 مع الترقيات الأخيرة48,49, المقابلة لمقاييس المكانية من ~ 1-600 Å. من ناحية أخرى، يتم حساب مقياس الوقت الذي يمكن الوصول إليه من زاوية السبق الكلية (أو المرحلة) التي يكتسبها النيوترون داخل لفائف السبق المغناطيسي، ووجد أن50: 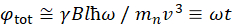 . في هذا التعبير، t هو الوقت فورييه
. في هذا التعبير، t هو الوقت فورييه 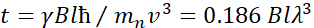 تعريفها بأنها ، حيث
تعريفها بأنها ، حيث 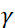 هي نسبة الدوران النيوتروني،
هي نسبة الدوران النيوتروني، 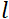 هو طول لفائف،
هو طول لفائف، 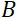 وقوة المجال المغناطيسي لللفائف. تجدر الإشارة إلى أن وقت فورييه هو كمية تعتمد بشكل صارم على هندسة الجهاز ، وقوة المجال المغناطيسي ، والطول الموجي النيوتروني. على سبيل المثال، باستخدام نيوترونات الطول الموجي
وقوة المجال المغناطيسي لللفائف. تجدر الإشارة إلى أن وقت فورييه هو كمية تعتمد بشكل صارم على هندسة الجهاز ، وقوة المجال المغناطيسي ، والطول الموجي النيوتروني. على سبيل المثال، باستخدام نيوترونات الطول الموجي 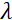 = 8 Å وإعدادات الصك = 1.2 متر و = 0.4 T، يتم حساب وقت فورييه ليكون t ~ 50 ns. تجريبيا، يتم ضبطها الوقت فورييه عن طريق تغيير التيار في لفائف قبل الخراج (أي قوة المجال المغناطيسي) أو باستخدام أطوال موجية نيوترونية مختلفة، مما أدى إلى جداول زمنية NSE نموذجية من ~ 1 PS إلى 100 ns. ومع ذلك، مكنت الترقيات الأخيرة في مطياف NSE الوصول إلى أوقات فورييه أطول، ما يصل إلى ~ 400 ns على مطياف J-NSE-Phoenix في هاينز ماير-ليبنيتز Zentrum51 ومطياف SNS-NSE في مختبر أوك ريدج الوطني48، وما يصل إلى ~ 1000 ns في مطياف NSE IN15 في معهد لو لانجفين (ILL)49.
= 8 Å وإعدادات الصك = 1.2 متر و = 0.4 T، يتم حساب وقت فورييه ليكون t ~ 50 ns. تجريبيا، يتم ضبطها الوقت فورييه عن طريق تغيير التيار في لفائف قبل الخراج (أي قوة المجال المغناطيسي) أو باستخدام أطوال موجية نيوترونية مختلفة، مما أدى إلى جداول زمنية NSE نموذجية من ~ 1 PS إلى 100 ns. ومع ذلك، مكنت الترقيات الأخيرة في مطياف NSE الوصول إلى أوقات فورييه أطول، ما يصل إلى ~ 400 ns على مطياف J-NSE-Phoenix في هاينز ماير-ليبنيتز Zentrum51 ومطياف SNS-NSE في مختبر أوك ريدج الوطني48، وما يصل إلى ~ 1000 ns في مطياف NSE IN15 في معهد لو لانجفين (ILL)49.
وإلى جانب الوصول المباشر إلى طول ديناميات الأغشية ونطاقها الزمني، فإن NSE لديها القدرات الكامنة في حساسية النظائر النيوترونية52. وعلى وجه التحديد، فإن قدرة النيوترونات على التفاعل بشكل مختلف مع نظائر الهيدروجين، وهو العنصر الأكثر وفرة في النظم البيولوجية، تؤدي إلى كثافة مختلفة لطول النيوترونات المتناثرة،34 أو NSLD (أي ما يعادل المؤشر البصري للانكسار50)،عندما يتم استبدال البروتيوم بالديوتريوم. وهذا يمكن من اتباع نهج يعرف باسم التباين التبايني، والذي يستخدم عادة لتسليط الضوء على ميزات غشاء محددة أو إخفاء الآخرين – ويشار إلى السيناريو الأخير على أنه مطابقة التباين. التطبيق المتكرر للتباين / المطابقة هو استبدال الماء (NSLD = -0.56 × 10-6 Å-2)بالماء الثقيل أو D2O (NSLD = 6 .4 × 10-6 Å-2)لتضخيم إشارة النيوترون من الأغشية الدهنية البروتية (NSLD ~ 0 × 10-6 Å-2). هذا النهج فعال للغاية في دراسات بنية الغشاء لأن اختراق D2O في منطقة مجموعة الرأس من الغشاء يسمح بتحديد دقيق لسماكة الغشاء (انظر الشكل 2A، اللوحة اليسرى) وموقع المجموعات الفرعية المختلفة للدهون عندما يتم تطبيق نماذج أكثر تطورا53،54. تسلط هذه الورقة الضوء على بعض الأمثلة على استخدام التباين في دراسات الديناميكيات الجماعية في الأغشية المحاكاة الحيوية وميزات الأغشية المختارة.
هنا ، تتجلى فعالية NSE في توفير رؤى فريدة في خصائص الأغشية الديناميكية والوظيفية من خلال أمثلة ملموسة لدراسات NSE على أنظمة أغشية الدهون النموذجية والوثيقة الصلة بيولوجيا مع التركيز على ديناميكيات مقياس المتوسط في الأغشية القائمة بذاتها ، في شكل تعليق ليبوسومي. بالنسبة لقياسات NSE لديناميكيات الأغشية داخل الطائرة ، يشار إلى القارئ إلى منشورات مخصصة حول التحليل الطيفي للنيوترونات الدورانية الصدى (GINSES)55و56 ودراسات أخرى لمداخن الأغشية متعددة اللاميلار المنحازة57و58و59و60.
للبساطة ، تسلط هذه الورقة الضوء على ثلاثة مخططات مختلفة من الديوتريشن الغشاء يتضح على تشكيل المجال مدروسة جيدا ، أو فصل المرحلة، نظام ثنائي الطبقات الدهون من 1،2-dimyristoyl-sn-glycero-3-فوسفوتشولين (DMPC) و 1،2-distearoyl-sn-glycero-3-فوسفوتشولين (DSPC) خلائط61،62. وتتميز الدهون اثنين من عدم تطابق في طول سلسلة الهيدروكربونات (14 الكربونات / الذيل في DMPC مقابل 18 الكربون / الذيل في DSPC) ودرجة الحرارة انتقال هلام السائل (TM، DMPC = 23 درجة مئوية مقابل Tم، DSPC = 55 درجة مئوية). وهذا يؤدي إلى فصل المرحلة الجانبية في DMPC: DSPC الأغشية في درجات الحرارة بين درجات الحرارة الانتقالية العليا والسفلى من الخليط63. يتم اختيار مخططات الديوتر التي تم النظر فيها هنا لإظهار الأنماط الديناميكية المختلفة التي يمكن الوصول إليها في قياسات NSE على الأغشية الدهنية ، وهي تقلبات الانحناء ، وتقلبات السماكة ، وتقلبات الانحناء / السماكة الانتقائية للمجالات الجانبية. يتم الإبلاغ عن جميع التراكيب الدهنية لDMPC: DSPC الطبقات الثنائية المعدة في جزء الخلد من 70:30، وذلك باستخدام المتغيرات المتاحة تجاريا بروتيات و perdeuterated من DMPC وDSPC. وتستند جميع خطوات إعداد العينة على 4 مل من تعليق الدهون، في D2O، مع تركيز الدهون من 50 ملغم / مل، لكتلة الدهون الإجمالية منتوت M= 200 ملغ لكل عينة.
