Het begrip van celmembranen en hun functie is de afgelopen decennia opmerkelijk geëvolueerd. De vroegere visie van celmembranen als passieve lipide-bilayers die celgrenzen definiëren en huismembraaneiwitten1 is geleidelijk veranderd in een dynamisch model waarin lipide-tweelagen een belangrijke rol spelen bij het reguleren van vitale biologische processen, waaronder cellulaire signalering, moleculaire uitwisseling en eiwitfunctie – om er maar een paar2,3,4,5,6te noemen . Dit besef dat celmembranen zeer dynamisch zijn en voortdurend een remodellering en moleculaire herverdeling ondergaan, heeft wetenschappelijke verkenningen aangedrongen die verder gaan dan evenwichtsstructuren van membranen7,8,9. Dienovereenkomstig zijn meerdere benaderingen ontwikkeld om de verschillende dynamische modi in biologische en bio-geïnspireerde lipidenmembranen te bestuderen. Tot op heden hebben de meeste van deze studies zich voornamelijk gericht op diffusieve moleculaire bewegingen10,11,12,13 en macroscopische vormschommelingen14,15,16, waardoor een aanzienlijke kloof is achtergelaten in het begrijpen van de intermediaire membraandynamiek, d.w.z. collectieve fluctuaties van lipidenassemblages bestaande uit enkele 10-100s lipidemoleculen. Deze dynamieken komen voor over lengteschalen van enkele tientallen tot enkele 100 Å en in de loop van de tijd schalen van sub-ns tot enkele honderden ns (zie figuur 1), hier aangeduid als mesoscopische schalen. Het is inderdaad op deze schalen dat belangrijke biologische activiteit plaatsvindt op membraanniveau17. Dit omvat virale ontluikende18, kanaal gating19, en membraan-eiwit interacties20. Het is ook belangrijk erop te wijzen dat het energielandschap van membraaneiwitten21,22 aantoont dat conformationele veranderingen in eiwitten — noodzakelijk voor hun regulerende rol — plaatsvinden in de tijdschalen23 van collectieve membraanschommelingen , waarbij het belang van mesoscopische dynamiek in de biologische functie van celmembranen en hun bio-geïnspireerde analogen verder wordt benadrukt20. Dit artikel richt zich op de twee primaire mesoscopische dynamische modi in lipidemembranen, namelijk buigschommelingen en dikteschommelingen.
De grootste uitdaging bij het direct onderzoeken van deze fluctuatiemodi is de moeilijkheid om tegelijkertijd toegang te krijgen tot hun ruimtelijke en temporele schalen met behulp van standaardspectroscopiemethoden. De andere uitdaging is dat directe contacttechnieken dezelfde fluctuaties kunnen beïnvloeden als die welke bedoeld zijn om16te meten . Dit wordt nog verergerd door de compositorische en structurele complexiteit van biologische membranen24,25, wat resulteert in niet-homogene membraankenmerken, waaronder lipidedomeinvorming26,27,28,29,30 en membraanasymmetrie31,32,33– die selectieve sondes vereisen om de dynamiek van verschillende membraankenmerken te begrijpen. Gelukkig kunnen deze uitdagingen worden overwonnen met niet-invasieve neutronenspectroscopiemethoden, zoals neutronenspinecho (NSE), die inherent toegang hebben tot de vereiste lengte- en tijdschalen, en verder studies van selectieve membraankenmerken mogelijk maken zonder hun fysisch-chemische omgeving te veranderen34. Inderdaad, in de afgelopen jaren is NSE spectroscopie geëvolueerd tot een unieke en krachtige sonde van collectieve membraandynamica35. Resultaten van NSE-studies over lipidemembranen hebben nieuwe inzichten opgeleverd in mechanische36,37 en visco-elastische38,39 eigenschappen van lipidemembranen en hebben nieuw licht geworpen op hun potentiële rol in biologische functie40,41.
De NSE-spectroscopietechniek is gebaseerd op een interferometrisch instrumentontwerp, voor het eerst voorgesteld door Mezei42, met behulp van een reeks spin-flippers en magnetische spoelen om de precessie van de neutronenspin te regelen terwijl neutronen het instrument doorkruisen. Het ontwerp berust op magnetische spiegeling van de magnetische veldelementen ten opzichte van de monsterpositie (figuur 1A). Dit houdt in dat bij gebrek aan energie-uitwisseling tussen het neutron en het monster, het neutron hetzelfde aantal spinprecessies uitvoert, in tegengestelde richting, in de eerste en tweede helft van het instrument (let op de π-flipper tussen de twee precessiespoelen). Als gevolg hiervan blijft de uiteindelijke spintoestand van het neutron ongewijzigd ten opzichte van de begintoestand – een fenomeen dat spin-echo wordt genoemd (zie transparant neutron in figuur 1A). Wanneer het neutron echter energetisch interageert met het monster, wijzigt de energie-uitwisseling het aantal spinprecessies in de tweede helft van het instrument, wat leidt tot een andere eindspintoestand (zie figuur 1A). Dit wordt experimenteel gedetecteerd als een verlies in polarisatie, zoals later in dit artikel zal worden getoond. Voor meer informatie over de NSE-techniek wordt de lezer verwezen naar speciale technische documenten42,43,44,45.
Hier wordt een vereenvoudigde beschrijving gepresenteerd om een ruwe schatting te geven van de lengte- en tijdschalen die toegankelijk zijn met NSE. De lengteschalen worden bepaald door het bereik van haalbare wavevectortransfers, Q = 4π sin θ/λ, waarbij 2θ de verstrooiingshoek is en λ de neutronengolflengte. Men kan zien dat Q wordt ingesteld door het golflengtebereik en de mate van rotatie van de tweede arm van de spectrometer (zie figuur 1A). Een typisch Q-bereikop NSE-spectrometers is ~0.02-2 Å-146,47en tot 0.01-4 Å-1 met recente upgrades48,49, overeenkomend met ruimtelijke schalen van ~1-600 Å. Aan de andere kant wordt de toegankelijke tijdschaal berekend op basis van de totale precessiehoek (of fase) die het neutron in de magnetische precessiespoelen heeft verkregen, en blijktdeze 50te zijn : 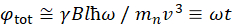 . In deze uitdrukking is t de Fouriertijd gedefinieerd als
. In deze uitdrukking is t de Fouriertijd gedefinieerd als 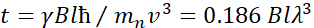 , waar is de
, waar is de 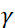 neutronen gyromagnetische verhouding,
neutronen gyromagnetische verhouding, 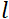 is de spoellengte, en
is de spoellengte, en 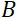 is de sterkte van het magnetisch veld van de spoel. Het is de moeite waard erop te wijzen dat de Fouriertijd een hoeveelheid is die strikt afhankelijk is van de instrumentgeometrie, de magnetische veldsterkte en de neutronengolflengte. Bijvoorbeeld, met behulp van neutronen van golflengte
is de sterkte van het magnetisch veld van de spoel. Het is de moeite waard erop te wijzen dat de Fouriertijd een hoeveelheid is die strikt afhankelijk is van de instrumentgeometrie, de magnetische veldsterkte en de neutronengolflengte. Bijvoorbeeld, met behulp van neutronen van golflengte 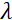 = 8 Å en instrumentinstellingen van = 1,2 m en = 0,4 T, wordt de Fouriertijd berekend op t ~ 50 ns. Experimenteel wordt de Fouriertijd afgestemd door de stroom in de precessiespoelen (d.w.z. magnetische veldsterkte) te veranderen of verschillende neutronengolflengten te gebruiken, wat resulteert in typische NSE-tijdschalen van ~ 1 ps tot 100 ns. Recente upgrades in NSE-spectrometers hebben echter toegang gegeven tot langere Fourier-tijden, tot ~ 400 ns op de J-NSE-Phoenix spectrometer op de Heinz Maier-Leibnitz Zentrum51 en de SNS-NSE spectrometer in Oak Ridge National Lab48, en tot ~ 1.000 ns op de IN15 NSE spectrometer in het Institut Laue-Langevin (ILL).
= 8 Å en instrumentinstellingen van = 1,2 m en = 0,4 T, wordt de Fouriertijd berekend op t ~ 50 ns. Experimenteel wordt de Fouriertijd afgestemd door de stroom in de precessiespoelen (d.w.z. magnetische veldsterkte) te veranderen of verschillende neutronengolflengten te gebruiken, wat resulteert in typische NSE-tijdschalen van ~ 1 ps tot 100 ns. Recente upgrades in NSE-spectrometers hebben echter toegang gegeven tot langere Fourier-tijden, tot ~ 400 ns op de J-NSE-Phoenix spectrometer op de Heinz Maier-Leibnitz Zentrum51 en de SNS-NSE spectrometer in Oak Ridge National Lab48, en tot ~ 1.000 ns op de IN15 NSE spectrometer in het Institut Laue-Langevin (ILL).
Naast directe toegang tot de lengte- en tijdschaal van membraandynamica heeft NSE de inherente mogelijkheden van neutronenisotopengevoeligheid52. Met name het vermogen van neutronen om verschillend te interageren met de isotopen van waterstof, het meest voorkomende element in biologische systemen, resulteert in een andere neutronenverstrooiingslengtedichtheid,34 of NSLD (het equivalent van de optische brekingsindex50), wanneer protium wordt vervangen door deuterium. Dit maakt een benadering mogelijk die bekend staat als contrastvariatie, die vaak wordt gebruikt om specifieke membraankenmerken te benadrukken of anderen te verbergen – het laatste scenario wordt contrastmatching genoemd. Een frequente toepassing van contrastvariatie/matching is de vervanging van water (NSLD = -0,56 × 10-6 Å-2) door zwaar water of D2O (NSLD = 6,4 × 10-6 Å-2) om het neutronensignaal van geprotieerde lipidemembranen (NSLD ~ 0 × 10-6 Å-2)te versterken. Deze aanpak is zeer effectief in studies van membraanstructuur omdat de penetratie van D2O in het hoofdgroepgebied van het membraan een nauwkeurige bepaling van de membraandikten mogelijk maakt (zie figuur 2A, linkerpaneel) en van de locatie van verschillende lipidesubgroepen wanneer meer geavanceerde modellen worden toegepast53,54. Dit artikel belicht enkele voorbeelden over het gebruik van contrastvariatie voor studies naar collectieve dynamiek in biomimetische membranen en bepaalde membraankenmerken.
Hier wordt de effectiviteit van NSE bij het bieden van unieke inzichten in dynamische en functionele membraaneigenschappen geïllustreerd aan de hand van tastbare voorbeelden van NSE-studies naar model- en biologisch relevante lipidenmembraansystemen met de nadruk op mesoschaaldynamiek in vrijstaande membranen, in de vorm van liposomale suspensuspensuspensussen. Voor NSE-metingen van membraandynamica in het vlak wordt de lezer verwezen naar speciale publicaties over beweidingsincidentie neutronenspin-echospectroscopie (GINSES)55,56 en andere studies van uitgelijnde multilamellar membraanstapels57,58,59,60.
Voor de eenvoud belicht dit artikel drie verschillende schema’s van membraandeuteratie geïllustreerd op een goed bestudeerd domeinvormend, of fasescheidend, lipidenbilayer-systeem van 1,2-dimyristoyl-sn-glycero-3-phosphocholine (DMPC) en 1,2-distearoyl-sn-glycero-3-phosphocholine (DSPC) mengsels61,62. De twee lipiden worden gekenmerkt door een mismatch in hun koolwaterstofketenlengte (14 carbons/tail in DMPC vs 18 carbons/tail in DSPC) en hun gel-fluid overgangstemperatuur (Tm, DMPC = 23 °C vs Tm, DSPC = 55 °C). Dit resulteert in laterale fasescheiding in DMPC:DSPC-membranen bij temperaturen tussen de bovenste en onderste overgangstemperaturen van het mengsel63. De hier overwogen deuteratieschema’s zijn gekozen om de verschillende dynamische modi aan te tonen die toegankelijk zijn in NSE-metingen op liposomale membranen, namelijk buigschommelingen, dikteschommelingen en selectieve buig-/dikteschommelingen van laterale domeinen. Alle lipidesamenstellingen worden gerapporteerd voor DMPC:DSPC-bilayers bereid met een molfractie van 70:30, met behulp van in de handel verkrijgbare protiated en perdeuterated varianten van DMPC en DSPC. Alle monstervoorbereidingsstappen zijn gebaseerd op 4 ml liposomale suspensie, in D2O, met een lipideconcentratie van 50 mg/ml, voor een totale lipidenmassa van Mtot = 200 mg per monster.
