For the analysis of embryonic development and maturation, bovine discs were used. To evaluate degeneration of the IVD, human samples were analyzed.
Human IVD tissue was obtained from patients undergoing surgery for lumbar disc degeneration, disc prolapse, or spinal trauma in the Department of Orthopaedic Surgery, University Hospital of Tübingen and the BG Trauma Centre Tübingen. Full ethical committee approval was obtained before the commencement of the study (project number 244/2013BO2). Written informed consent was received from all patients before participation. The methods were carried out in accordance with the approved guidelines.
Bovine tissue was obtained from the Bavarian State Office for Health and Food Safety/Oberschleißheim and from a rendering plant in Warthausen (Germany). Local and veterinary authorities' approval was received for tissue from dead animals.
1. Sample harvest
- Human IVD tissue: Place intraoperatively-obtained IVD samples immediately in Dulbecco's Modified Eagle Medium (DMEM) with 2% (v/v) of penicillin-streptomycin and 1.2% of (v/v) amphotericin B. Store at 4 °C until further processing. Process the tissue within 48 h. Alternatively, store the tissues at -20 °C for several weeks.
- Bovine IVD tissue: Ensure to harvest the tissue from the animals within 24 h after death.
Resect the bovine discs with the surrounding vertebrae from the dead animals en-bloc. Transport the frozen tissue on dry ice and store at -20 °C until further processing.
NOTE: If only fluorescence analyses are intended and no further biochemical quantification methods such as ELISA or PCR are planned, perform the tissue fixation as explained below. This allows to keep the tissue longer in storage before it needs to be processed. To prevent deterioration of the tissue matrix, perform fixation within 48 h after harvest unless the tissue is frozen directly.
2. Sample preparation
- Thaw the frozen tissue at room-temperature. Process the tissue as soon as no ice-crystals can be felt any more upon digital gentle compression of the tissue.
NOTE: Perform preparation of the tissue in DMEM in a Petri dish. - Identify the origin of the human IVD tissue (anulus fibrosus, intermediate zone, nucleus pulposus, or cartilaginous endplate) based on macroscopic properties such as collagen density and orientation.
- Take the motion segment consisting of the bovine IVD disc with its two adjacent vertebrae and dissect the disc as a whole from the subchondral bone using a surgical blade (blade number 15).
- Use two anatomic forceps to flip the disc to reach areas more centered. Perform the dissection. Ensure to resect the nucleus pulposus last as it is much thinner than the anulus, prone to tear, and it does not come off in a defined fashion easily.
- Identify the different areas of the cartilage.
- Cut out the area of interest from the whole disc using a surgical blade (blade number 20 or 22). Alternatively, use a cryotome blade.
NOTE: As bovine discs come en-bloc as a part of the spine, the discs can be prepared in-toto. This makes correct identification of the different types of cartilage much easier. When dissecting the disc in the fashion described above, the cartilaginous endplate remains on the vertebrae. If this area shall be investigated, it is best taken off the underlying bone with a chisel working in a slightly bent tangential direction.
- In case that embryos are of a crown-rump length of smaller than 20 cm, ensure to process the embryos in toto to preserve the tissue architecture of the IVD. Do not perform any dissection of the vertebrae in these cases.
- Once the disc has been resected in-toto, identify the different areas of the cartilage.
- Perform the decalcification in ethylenediaminetetraacetic acid (EDTA) (20% (w/v); pH = 7.4) at room temperature. Choose the volume depending on the sample size – the entire tissue must be well covered with EDTA.
- Change the decalcification solution daily which can go for up to 5 days depending on the size of the tissue.
- Verify that the decalcification is successful with a 20 Gauge needle that penetrates the vertebrae without notable resistance.
NOTE: The daily change of decalcification solution is important to prevent the chelate binder EDTA from saturation to maintain reaction effectivity. It also prevents bacterial colonization.
3. Grading of sample age, integrity, and degeneration
- Classify the human disc tissue into one of the five following categories with the help of clinical information as well as X-ray and magnetic resonance imaging18 (Figure 3).
NOTE: Category I: To serve as near-healthy sample use anulus without any radiological sign of IVD degeneration derived from acute spinal trauma.
Category II: To illustrate a situation of acute inflammation with beginning degeneration use tissue from the intermediate zone from patients with clinical symptoms with a duration of less than 4 weeks.
Category III: To describe a situation where the inflammatory reaction had already had time to affect the tissue and cells take tissue from patients who were operated for a nuclear prolapse but with a symptom duration exceeding 4 weeks.
Category IV: For moderate disc degeneration select anulus obtained from surgery with interbody fusion for degenerative disc disease with a Pfirrmann score of 3 or 4 in magnetic resonance imaging19.
Category V: For end stage degeneration process anulus obtained from surgery with interbody fusion for degenerative disc disease with a Pfirrmann score of 5. - Classify the bovine tissue based on the developmental stage/age of the animal into one of the eight categories, as displayed in Table 1.
- Calculate the gestation age on crown-rump length of the embryos based on the formula suggested by Keller:
Gestation age in months =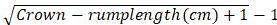
NOTE: Animals in the first 4 weeks of gestation present with a crown-rump length of 0.8-2.2 cm20.
- Calculate the gestation age on crown-rump length of the embryos based on the formula suggested by Keller:
4. Tissue fixation
- Fixate the samples in 10x the volume of the sample of 4% (w/v) formaldehyde solution in phosphate buffered saline (PBS) over night at 4 °C.
NOTE: The formaldehyde solution penetrates tissue at a rate of about 1 mm/hour from each direction. For very small or very big samples an adjustment of the exposure time could be necessary. - Store the tissue in PBS at 4 °C until further processing.
5. Histologic sectioning
- Embed the samples in water-soluble embedding medium on the cryotome knob.
- Place the tissue onto the knob such that either an axial plane is generated or a plane that cuts the collagen type I lamellae perpendicular (e.g., a median sagittal sectioning plane).
NOTE: The tissue has to be fully covered by the embedding medium.
- Place the tissue onto the knob such that either an axial plane is generated or a plane that cuts the collagen type I lamellae perpendicular (e.g., a median sagittal sectioning plane).
- Section the embedded tissue at a thickness of 70 µm in human samples and 40 µm in bovine samples using a standard cryotome.
NOTE: The difference in section thickness is due to the difference in tissue integrity between the intact bovine disc and the highly degenerate human tissue. - Collect the sections on a glass slide.
- Encircle the tissue sections with a hydrophobic pen.
- Rinse the sections 3 times with phosphate-buffered saline (PBS) to remove the water-soluble embedding medium.
6. Fluorescence staining
- Add 60 µL of 1% (v/v) of DAPI (Exmax 358 nm, Emmax 461 nm) as well as 1% (v/v) of Actin Tracking staining (Exmax 540 nm, Emmax 565 nm) in PBS and incubate for 5 min at room temperature.
NOTE: The staining protocol described here is to visualize the nucleus using DAPI nuclear staining (blue) and the cytoplasm using Actin Tracking Stain (red). The IVD has a strong autofluorescence due to the collagen fibers in the green channel. The amount of fluid added to the sections in this protocol is intended for sections of a size of about 5 mm x 5 mm. For larger sections, this amount needs to be increased accordingly. Perform all the works in rooms without direct sunlight exposure and with dimmed lights to prevent bleaching the dye. - Remove the staining fluid with a pipette and wash three times with 60 µL of PBS each time.
- Add a suitable mounting medium and cover the sections with a cover slip.
NOTE: Ensure that there are no air bubbles entrapped when adding the cover slip. This is best done by starting contact of the slip with the slide on one rim and then let the slip come down slowly.
7. Microscopic imaging and processing
- Place a slide with a stained section on the sample holder of the microscope.
NOTE: Due to the dense collagen type I network of the IVD, the scattered light makes the tissue difficult to visualize using conventional fluorescence microscopy. One way to address this problem is to perform optical sectioning using structured illumination. This also allows to render a three-dimensional projection of the entire specimen in both channels (blue and red). This is best done using the structured illumination setting and the Mosaic-mode with a 10x magnification objective lens to obtain an overview of the sample as well as 3D reconstructions of individual patterns. - Start the structured illumination device.
- Perform single field-of-view imaging with a suitable fluorescence microscope, fluorescence filters and adequate illumination.
NOTE: Adjust the exposure time for all the filters used in order to standardize the imaging acquisition. To get an accurate representation of the specimen at a higher resolution image the sections with a higher magnification (e.g., 20x objective). - Postprocess the pictures by optimizing the intensity and brightness using an image optimization software compatible with the fluorescence microscope.
- Perform single field-of-view imaging with a suitable fluorescence microscope, fluorescence filters and adequate illumination.
- To visualize the section as a whole use the mosaic imaging technique
- Open the acquisition settings (press on 6D- Acquisition) from the toolbar panel.
- Adjust the mosaic settings (in MosaiX Register) and define the number of columns and rows of field-of-view images that shall later be merged to one overview image.
- Press Setup and adjust the focus correction of individual tiles.
NOTE: It is almost impossible for a large tissue section to have the whole tissue surface in a single focus plane. Image tiles at different focal levels can be taken by 'MosaiX Acquisition'. - To start the acquisition of the image tiles, press Start.
- Stitch the imaged tiles by using the stitching function (Stitching button) with 20% overlap incorporated in the software.
- Postprocess the pictures by optimizing the intensity and brightness using an image optimization software compatible with the fluorescence microscope.
- To analyze the spatial chondrocyte organization, use the 3D function incorporated in the software.
- Adjust the z-stack settings. Define the scanning parameters: define the start and stop positions in the z-axis and the slice distance by activating Start/Stop button.
NOTE: The software automatically calculates the number of slices. - To start acquisition of the image z-stacks, press Start.
- Postprocess the pictures by optimizing the intensity and brightness using an image optimization software compatible with the fluorescence microscope.
- Adjust the z-stack settings. Define the scanning parameters: define the start and stop positions in the z-axis and the slice distance by activating Start/Stop button.
- Export the pictures with a file format compatible with the image processing software.
NOTE: Export the 3D reconstructions as individual images or/and as an interactive 3D model or in a video format.
8. Cellular pattern identification and density assessment
- Open the exported mosaic pictures of the entire tissue section in an appropriate image processing program.
- Define the areas subjected to cell density assessment by defining regions of interest of 500 µm x 500 µm in the pictures.
NOTE: All tiles that do not have an adequate image quality are excluded from analysis. - Identify individual cellular patterns.
NOTE: Single cells are defined as individual cells – fully encapsulated within the adjacent matrix. Pairs are defined as two adjacent cells in close proximity (<25 µm) whereby the cells are interconnected through their matrixes (see Figure 2). String-formations are at least three chondrocytes aligned in line (from the middle of the nuclei <25 µm). These cells are encompassed by an intact matrix and matrix interconnections can be seen between each cell. Clusters represent multiple cells that are located in direct proximity to each other (<25 µm) and are encapsulated in a large lacuna devoid of matrix. - Use a cell count plug-in for the quantitative analysis of the cellular patterns.
NOTE: The cytoplasm staining represents a verification method to identify the different spatial patterns. - Calculate the cell density by dividing the counted cells by the size of the chosen region of interest.
Using mosaic images, the architecture of the IVD with its dense collagen fiber network in the anulus and the softer nucleus can clearly be recognized (Figure 4). A continuous decrease in cellular density can be observed during embryonic development (Figure 5). While in the early stages of IVD development a cell density of 11,435 cells/mm² in the bovine anulus fibrosus and 17,426 cells/mm² in the bovine nucleus pulposus can be found, these numbers decrease rapidly to 1,011 cells/mm² (bovine anulus fibrosus) and 488 cells/mm² (bovine nucleus pulposus) until birth. In the adult cattle 71 cells/mm² (anulus fibrosus) resp. 106 cells/mm² (nucleus pulposus) are seen (Figure 6 A-B). Using bi-channel imaging with the Apotome allows to visualize the 3D architecture of the spatial patterns (Figure 7).
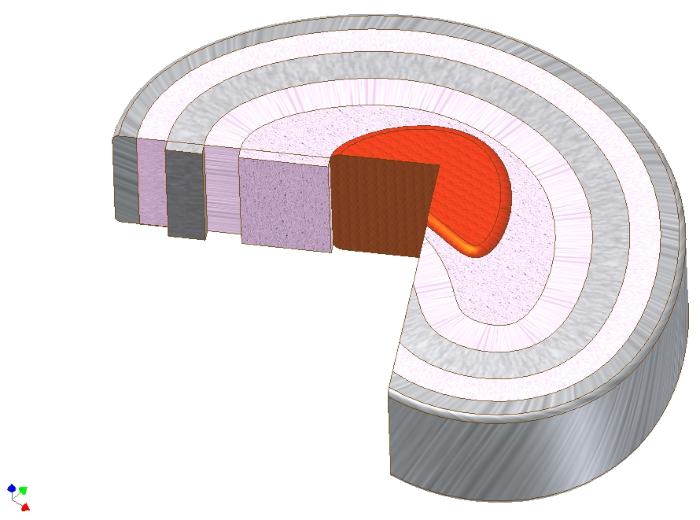
Figure 1. Macroscopic anatomy of the intervertebral disc. Schematic drawing of the intervertebral disc showing the nucleus pulposus (red), directly around it the intermediate zone (pink), and then in circular layers around it the anulus fibrosus. Note the ply-angle direction of the collagen type I fibers in the anulus fibrosus. The axial load to the nucleus can thus be translated to axial tensile forces of the collagen fibers. Please click here to view a larger version of this figure.
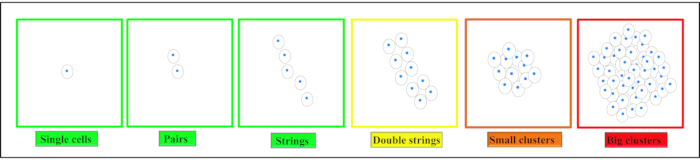
Figure 2. Schematic illustration of different spatial organizational patterns of chondrocytes. Depending on the tissue chondrocytes are found as single cells, pairs or strings in healthy cartilage. With beginning degeneration these patterns change to form double strings, small clusters and then big clusters in end-stage degeneration. This figure has been modified from Danalache, M. et al.21. Please click here to view a larger version of this figure.
Figure 3. Different conditions of the intervertebral disc (IVD). (A–D') Sagittal T2-weighted magnetic resonance imaging of the human lumbar spine (A–D), with the magnified motion segment L4/L5 (A'–D'). The ventral side of the patient faces left, the dorsal side faces right with the spinal canal with its white signal for the cerebrospinal fluid. (A,A') Intact IVD with the anulus displayed with a hypointense (black) signal due to the high collagen type I content and the nucleus displayed much brighter (hyperintense) due to the high content of water-binding proteoglycans (Pfirrmann grade19 I). (B,B') Beginning IVD degeneration with a loss of the water signal from the nucleus pulposus and where the distinction between the anulus and nucleus is lost (Pfirrmann grade19 IV). (C,C') Acute nuclear prolapse with still a prominent water signal in the area of the nucleus indicative for an otherwise intact IVD and the disc tissue protruding dorsally into the spinal canal. (D,D') Advanced disc degeneration with a largely destroyed IVD with a complete loss of the water signal within the disc, ventral and dorsal spondylophyte formation and subchondral sclerosis of the vertebrae corresponding to a Pfirrmann grade19 of V. Please click here to view a larger version of this figure.
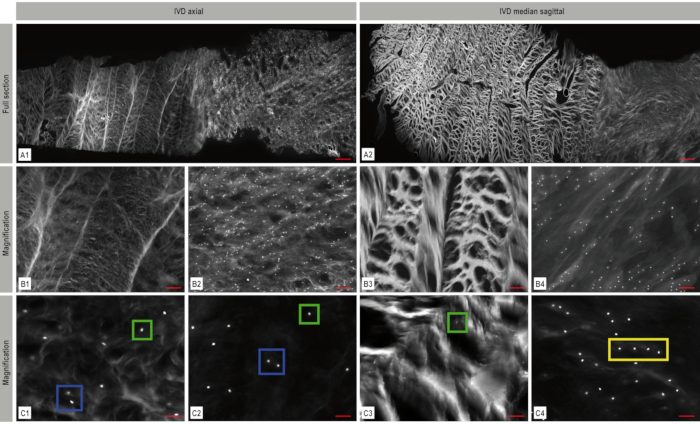
Figure 4. Mosaic fluorescence imaging of the intervertebral disc (IVD). The architecture of the IVD with its dense collagen fiber network in the anulus fibrosus and the softer nucleus can clearly be recognized. DAPI-nuclear staining (white) in the axial (A1) and sagittal (A2) plane shows the cell distribution and arrangement within the IVD. Magnified representative areas from these mosaic images are displayed in B1–B4 and C1–C4 illustrating the spatial chondrocyte organization – in this case single cells (green box), pairs (blue box) and strings (yellow box). A: scale bar 1,000 µm, B1–B4: scale bar 200 µm, C1–C4: scale bar: 50 µm. This figure has been modified from Bonnaire, F. C. et al.22. Please click here to view a larger version of this figure.

Figure 5. Different developmental and maturation stages of bovine anulus fibrosus and human degenerating anulus fibrosus. DAPI nuclear staining. The mosaic images show a whole early embryonic disc in an axial section (A1) and representative images of the bovine anulus during embryonic development, maturation, and beginning degeneration (A2–A8). (B1–B3) Anulus from human IVDs was obtained intraoperatively. A continuous decrease in cellular density can be observed during embryonic disc development. A higher spatial organization cellular pattern seems to be present especially around birth. In the adult human disc during degeneration, the cellular density increases again and increasing cluster formation can be observed. Scale bar 100 µm. Wog: weeks of gestation. This figure has been modified from Bonnaire, F. C. et al.22. Please click here to view a larger version of this figure.
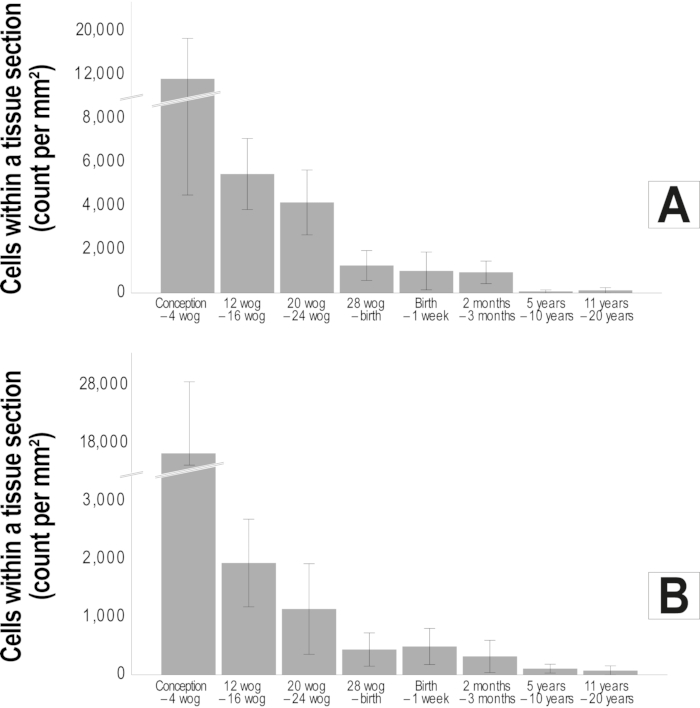
Figure 6. Reduction in cellular density throughout development and maturation of the bovine intervertebral disc. The mean (standard deviation) cell count per mm² is illustrated by bar diagrams for the bovine anulus fibrosus (A) and the bovine nucleus pulposus (B). A clear reduction of cellular density can be observed especially during the embryonic period which continues to a lesser extent at least until full maturation of the disc (n=72). Wog: weeks of gestation. This figure has been modified from Bonnaire, F. C. et al.22. Please click here to view a larger version of this figure.
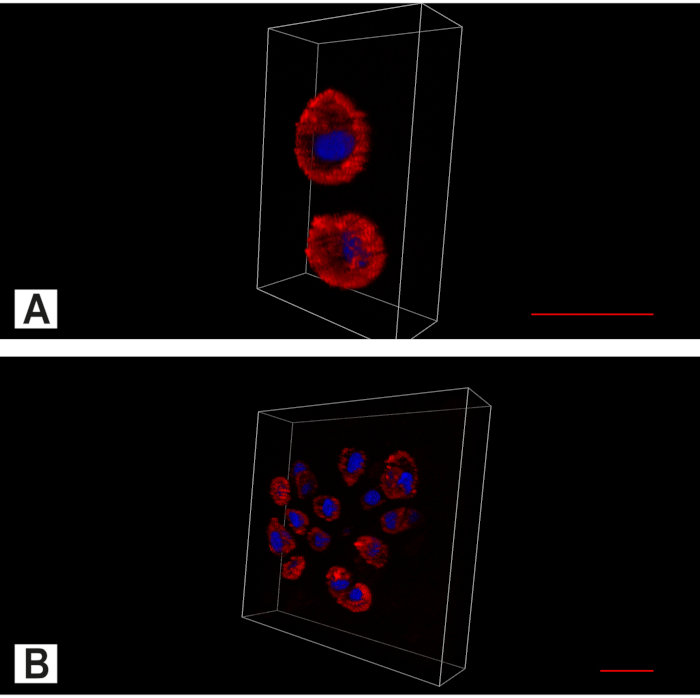
Figure 7. Apotome imaging of the intervertebral disc (IVD). Bi-channel image showing the cytoplasm (red, Actin staining) and the nucleus (blue, DAPI nuclear staining). (A) In the intact IVD, in addition to the predominant spatial pattern of single chondrocytes, pairs are also found. (B) In degenerated anulus cell clusters can be found. Scale bar 20 µm. Please click here to view a larger version of this figure.
Video 1. Apotome imaging of a pair in the intervertebral disc (IVD) as a 3D Model. Bi-channel images showing the cytoplasm (red, Actin staining) and the nucleus (blue, DAPI nuclear staining) of a pair. Please click here to download this Video.
Video 2. Apotome imaging of a cluster in the intervertebral disc (IVD) as a 3D Model. Bi-channel images showing the cytoplasm (red, Actin staining) and the nucleus (blue, DAPI nuclear staining) of a cluster. Please click here to download this Video.
Table 1: Bovine embryonic development, maturation, and growth after birth with its different milestones. Wog – weeks of gestation. Gestation period ca. 283 days, natural life-expectancy 20-25 years20,23,24. Please click here to download this Table.
