Un obstacle majeur dans l’étude des maladies fibrotiques est le manque de modèles de tissus 3D humains représentatifs qui fournissent un aperçu du comportement des fibroblastes et de leurs dérivés pathologiques. Pour étudier les processus fibrotiques, les systèmes de culture 2D standard sont sous-optimaux puisque les fibroblastes isolés se transdifférencient rapidement en myofibroblastes exprimant l’actine musculaire α lisse (SMA) lorsqu’ils sont cultivés sur des substrats 2D non conformes1,2,3. Ainsi, les fibroblastes de la culture 2D standard ne reflètent pas un phénotype tissulaire « sain » régulier3,4,5,6. Des cultures sur substrats souples ont été introduites pour simuler des environnements tissulaires non fibrotiques (10 kPa) et fibrotiques (35 kPa)7, mais ceux-ci n’ont pas la troisième dimension, ce qui est très important en ce qui concerne la physiopathologie. L’ingénierie tissulaire offre la possibilité de surmonter cette limitation en permettant la culture de fibroblastes dans un contexte défini et expérimentalement accordable de matrice extracellulaire (ECM), par exemple, par des altérations de la cellularité, de la composition ecm et de la concentration ECM, qui peuvent toutes déterminer la biomécanique tissulaire.
Divers modèles 3D ont été générés à l’aide de fibroblastes. Les disques flottants et les microsphères ont été parmi les premiers et démontrent que le collagène est remodelé et compacté en fonction du temps. Les fibroblastes exercent des forces de traction sur les fibrilles de collagène, un processus qui peut être facilité par l’ajout d’agents pro-fibrotiques tels que le facteur de croissance transformant bêta 1 (TGF-β1)8,9,10,11,12,13,14,15,16. Cependant, les cultures flottant librement ne permettent pas la charge externe contrôlée et constituent donc des modèles de rétrécissement ou de compactage continus. Les tissus modifiés en forme de feuille ont ouvert la possibilité d’étudier la régulation homéostatique des propriétés biomécaniques des tissus, notamment par des tests de déformation uni, bi, multiaxiale ou cyclique.17,18,19,20. Ces modèles ont été utilisés, par exemple, pour démontrer l’influence du nombre de cellules sur la rigidité tissulaire, qui s’est avérée être en corrélation positive avec l’intégrité du cytosquelette et la contractilité du cytosquelette d’actomyosine.18,19. Cependant, il est important de noter que les conversions force-déformation sont compliquées par la déformation tissulaire non uniforme autour des points de serrage des transducteurs de force et des points d’ancrage. Cette limitation inhérente peut être contournée par des tissus en forme d’os de chien ou d’anneau, offrant une certaine application tissulaire aux points d’ancrage.21,22,23. Les tissus en forme d’anneau peuvent être préparés en distribuant un hydrogel de collagène cellulaire dans des moules en forme d’anneau. Au fur et à mesure que l’hydrogel se compacte, un tissu se forme autour de la tige interne incompressible du moule, ce qui offre une résistance à la contraction ultérieure des tissus.24,25,26,27. Après un compactage initial et généralement maximal, les tissus peuvent également être transférés à des entretoises réglables pour restreindre davantage l’ECT circulaire à une longueur de tissu définie.3,24,25,26,27,28,29,30. Les propriétés biophysiques peuvent être évaluées dans des dispositifs de contrainte de déformation horizontaux ou verticaux standard avec des capteurs de pesage appropriés sous déformation unidirectionnelle ou dynamique3. Comme les tissus ont une structure circulaire largement uniforme et peuvent être maintenus sur des barres / crochets (points d’ancrage et / ou transducteurs de force), bien que ceux-ci puissent encore entourer des zones de compression autour des barres de chargement, ce format permet une variation de déformation plus uniforme par rapport au serrage3. De plus, les tissus ancrés provoquent une forme de cellule bipolaire, et les cellules s’adaptent aux forces tissulaires en s’allongeant le long des lignes de force favorisant la traction anisotrope31,32,33,34,35,36. Nous avons déjà appliqué de l’ECT en forme d’anneau à partir de fibroblastes cardiaques (FK) de rats et d’humains autour d’un seul pôle rigide dans des expériences de contrainte fonctionnelle et effectué des études de gain et de perte de fonction en utilisant des fibroblastes transductés viralement24,25,26 et études pharmacologiques37. De plus, nous avons pu identifier les différences entre les sexes dans la fibrose médiée par la mucoviscidose dans le modèle ECT27.
Le protocole suivant pour la génération d’ECT humain, illustré par la mucoviscidose humaine primaire obtenue sous forme de mucoviscidose cryoconservée auprès de fournisseurs commerciaux (voir Tableau des matériaux), combine les avantages des tissus en forme d’anneau avec un moyen simple et rapide de produire des tissus macroscopiques pour une plate-forme de 48 puits conçue pour des tests parallèles à haute teneur.
Il est important de noter que le modèle ECT ne se limite pas à un type spécifique de fibroblastes, avec l’utilisation documentée dans l’étude d’autres fibroblastes, par exemple les fibroblastes de la peau38,39. De plus, les fibroblastes issus des biopsies des patients fonctionnent tout aussi bien, et le choix des fibroblastes dépend en fin de compte de la question scientifique à aborder.
La plate-forme utilisée pour la génération d’ECT décrite dans ce protocole est une plaque de culture cellulaire/tissulaire 3D de 48 puits disponible dans le commerce (Figure 1A). Les méthodes de préparation, de culture et de surveillance de la formation et du fonctionnement de l’ECT sous une géométrie et une charge mécanique définies à l’aide de la plaque de 48 puits sont décrites. Les ECT formés sont maintenus par des poteaux flexibles intégrés et la charge mécanique peut être ajustée en fonction de l’objectif final en utilisant des poteaux de dureté différente (valeur Shore A 36-89), influençant leurs rigidités en flexion. Les poteaux avec un rivage Une valeur de 46 sont recommandés. Le protocole est, en outre, compatible avec un moule circulaire personnalisé décrit précédemment, où l’ECT est maintenu autour d’une seule tige rigide37. Les dimensions de ce moule sont données à la figure 1B.
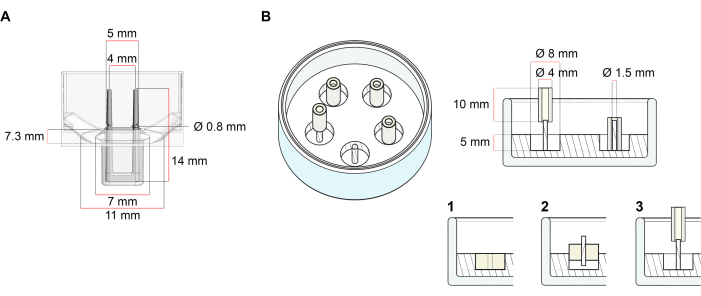
Figure 1 : Représentation schématique des moules de coulée. (A) Dessin technique et dimensions d’un moule de coulée avec deux pôles flexibles. Le moule comprend une circonférence intérieure délimitée par une paroi courte qui maintient des doubles pôles de retenue au corps principal du moule. Les pôles flexibles ont une distance horizontale libre les uns par rapport aux autres et sont connectés à la base. Le moule permet un volume de coulée de 180 μL. Le puits de chaque moule permet une capacité de volume d’au moins 600 μL de milieux de culture. Différentes compositions de matériaux peuvent être utilisées pour produire des pôles avec des rigidités spécifiques (par exemple, TM5MED-TM9MED). (B) Dessin technique et dimensions d’un moule en forme d’anneau avec une seule tige rigide. Il s’agit d’un moule alternatif avec une géométrie et un environnement mécanique distincts, qui peut être utilisé avec le protocole de coulée ECT37. La méthode d’assemblage du moule en forme d’anneau a été adaptée à partir de formats plus grands publiés28,41. En bref, la méthode comprend (1) l’impression d’entretoises de moulage en polytétrafluoroéthylène (PTFE) (8 mm de diamètre) dans du polydiméthylsiloxane (PDMS, silicone) versé dans des boîtes en verre (diamètre 60 mm), et (2) la fixation d’un support de poteau PDMS (1,5 mm de diamètre) concentriquement à l’intérieur de la cavité creuse formée, qui sert à (3) maintenir un pôle amovible (tube en silicone de 4 mm de diamètre). L’espace creux résultant permet 180 μL de volume de coulée. Chaque plat en verre peut contenir plusieurs moules imprimés (illustrés de manière exemplaire avec 5 moules) et a une capacité allant jusqu’à 5 mL de milieu de culture. Veuillez cliquer ici pour voir une version agrandie de cette figure.
