Un obstáculo importante en el estudio de las enfermedades fibróticas es la falta de modelos representativos de tejidos humanos en 3D que proporcionen información sobre el comportamiento de los fibroblastos y sus derivados patológicos. Para estudiar los procesos fibróticos, los sistemas de cultivo 2D estándar son subóptimos, ya que los fibroblastos aislados se transdiferencian rápidamente en miofibroblastos que expresan actina del músculo α liso (AME) cuando se cultivan en sustratos 2D no conformes1,2,3. Por lo tanto, los fibroblastos en el cultivo 2D estándar no reflejan un fenotipo tisular “sano” regular3,4,5,6. Se han introducido cultivos sobre sustratos flexibles para simular ambientes tisulares no fibróticos (10 kPa) y fibróticos (35 kPa)7, pero estos carecen de la tercera dimensión, que es muy importante con respecto a la fisiopatología. La ingeniería de tejidos brinda la oportunidad de superar esta limitación al permitir el cultivo de fibroblastos en un contexto de matriz extracelular (ECM) definido y experimentalmente sintonizable, por ejemplo, por alteraciones en la celularidad, la composición de ECM y la concentración de ECM, todo lo cual puede determinar la biomecánica del tejido.
Se han generado varios modelos 3D utilizando fibroblastos. Los discos flotantes y las microesferas estuvieron entre los primeros y demuestran que el colágeno se remodela y compacta de una manera dependiente del tiempo. Los fibroblastos ejercen fuerzas de tracción sobre las fibrillas de colágeno, un proceso que puede facilitarse mediante la adición de agentes profibróticos como el factor de crecimiento transformador beta 1 (TGF-β1)8,9,10,11,12,13,14,15,16. Sin embargo, los cultivos que flotan libremente no permiten la carga externa controlada y, por lo tanto, constituyen modelos que se encogen o compactan continuamente. Los tejidos diseñados en forma de lámina abrieron la posibilidad de estudiar la regulación homeostática de las propiedades biomecánicas de los tejidos, es decir, a través de pruebas de deformación uni, bi, multiaxial o cíclica.17,18,19,20. Estos modelos se han utilizado, por ejemplo, para demostrar la influencia del número de células en la rigidez del tejido, que se encontró que se correlaciona positivamente con la integridad del citoesqueleto y la contractilidad del citoesqueleto de actomiosina.18,19. Sin embargo, es importante tener en cuenta que las conversiones de fuerza a deformación se complican por la deformación no uniforme del tejido alrededor de los puntos de sujeción de los transductores de fuerza y los puntos de anclaje. Esta limitación inherente puede ser evitada por tejidos de hueso de perro o en forma de anillo, ofreciendo cierta aplicación de tejidos en los puntos de anclaje.21,22,23. Los tejidos en forma de anillo se pueden preparar distribuyendo un hidrogel de colágeno celular en moldes en forma de anillo. A medida que el hidrogel se compacta, se forma un tejido alrededor de la varilla interna incompresible del molde, que ofrece resistencia para una mayor contracción del tejido.24,25,26,27. Después de la compactación inicial y típicamente máxima, los tejidos también pueden transferirse a espaciadores ajustables para restringir aún más la TEC circular a una longitud de tejido definida.3,24,25,26,27,28,29,30. Las propiedades biofísicas pueden evaluarse en dispositivos estándar de tensión por deformación horizontal o vertical con células de carga adecuadas bajo deformación unidireccional o dinámica3. Como los tejidos tienen una estructura circular en gran medida uniforme y se pueden sostener en barras / ganchos (puntos de anclaje y / o transductores de fuerza), aunque estos aún pueden encerrar áreas de compresión alrededor de las barras de carga, este formato permite una variación de deformación más uniforme en comparación con el sujeción.3. Además, los tejidos anclados provocan una forma de célula bipolar, y las células se adaptan a las fuerzas del tejido por elongación a lo largo de las líneas de fuerza que promueven la tracción anisotrópica.31,32,33,34,35,36. Anteriormente hemos aplicado TEC en forma de anillo de fibroblastos cardíacos (FQ) de rata y humanos alrededor de un solo polo rígido en experimentos funcionales de tensión y tensión y hemos realizado estudios de ganancia y pérdida de función mediante el uso de fibroblastos transducidos viralmente.24,25,26 y estudios farmacológicos37. Además, pudimos identificar diferencias de sexo en la fibrosis mediada por FQ en el modelo de TEC27.
El siguiente protocolo para la generación de TEC humana, ejemplificado con la FQ humana primaria obtenida como FQ criopreservada de proveedores comerciales (ver Tabla de Materiales), combina las ventajas de los tejidos en forma de anillo con una forma fácil y rápida de producir tejidos macroscópicos para una plataforma de 48 pocillos diseñada para pruebas paralelas de alto contenido.
Es importante destacar que el modelo TEC no se restringe a un tipo específico de fibroblastos, con el uso documentado en la investigación de otros fibroblastos, por ejemplo, fibroblastos cutáneos38,39. Además, los fibroblastos de las biopsias del paciente funcionan igualmente bien, y la elección de los fibroblastos depende en última instancia de la cuestión científica que se debe abordar.
La plataforma utilizada para la generación de TEC descrita en este protocolo es una placa de cultivo de células/tejidos 3D de 48 pocillos disponible comercialmente (Figura 1A). Se describen los métodos para la preparación, el cultivo y el monitoreo de la formación y función de ECT bajo una geometría definida y carga mecánica con la ayuda de la placa de 48 pocillos. Los ECT formados se sujetan mediante postes flexibles integrados y la carga mecánica se puede ajustar de acuerdo con el propósito final mediante el uso de postes con diferente dureza (valor Shore A 36-89), influyendo en sus rigideces de flexión. Se recomiendan los polos con una orilla Un valor de 46. El protocolo es, además, compatible con un molde circular personalizado descrito anteriormente, donde el ECT se mantiene alrededor de una sola varilla rígida37. Las dimensiones de este molde se dan en la Figura 1B.
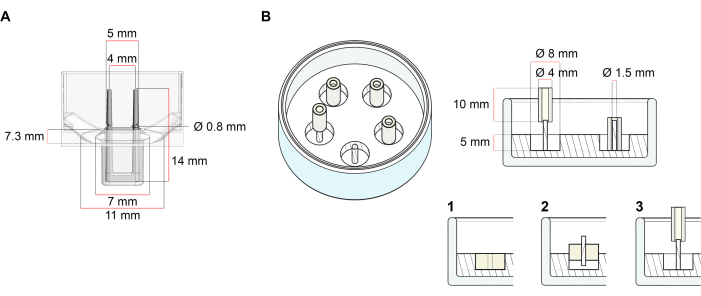
Figura 1: Representación esquemática de moldes de fundición. (A) Plano técnico y dimensiones de un molde de fundición con dos postes flexibles. El molde comprende una circunferencia interior delimitada por una pared corta que sostiene postes de retención dobles en el cuerpo principal del molde. Los polos flexibles tienen una distancia horizontal libre entre sí y están conectados en la base. El molde permite un volumen de fundición de 180 μL. El pozo de cada molde permite una capacidad de volumen de al menos 600 μL de medios de cultivo. Se pueden utilizar diferentes composiciones de materiales para producir postes con rigideces específicas (por ejemplo, TM5MED-TM9MED). (B) Dibujo técnico y dimensiones de un molde en forma de anillo con una sola varilla rígida. Se trata de un molde alternativo con geometría y entorno mecánico distintos, que se puede utilizar con el protocolo de fundición ECT37. El método de ensamblaje de moldes en forma de anillo se adaptó de formatos más grandes publicados28,41. En resumen, el método incluye (1) la impresión de espaciadores de moldeo de politetrafluoroetileno (PTFE) (8 mm de diámetro) en polidimetilsiloxano (PDMS, silicona) vertidos en platos de vidrio (diámetro 60 mm), y (2) la fijación de un soporte de poste PDMS (1,5 mm de diámetro) concéntricamente dentro de la cavidad hueca formada, que sirve para (3) sostener un poste extraíble (tubo de silicona de 4 mm de diámetro). El espacio hueco resultante permite 180 μL de volumen de fundición. Cada plato de vidrio puede contener múltiples moldes impresos (se muestra ejemplarmente con 5 moldes) y tiene la capacidad de hasta 5 ml de medio de cultivo. Haga clic aquí para ver una versión más grande de esta figura.
