Les chaperons moléculaires contribuent à l’homéostasie des protéines, y compris le repliement, le transport et la dégradation des protéines. Ils régulent plusieurs processus cellulaires et sont liés à de nombreuses maladies telles que le cancer et les maladies neurodégénératives1. La protéine de choc thermique 90 (Hsp90) est l’un des chaperons les plus importants dont la fonction dépend des changements conformationnels entraînés par l’hydrolyse de l’ATP et la liaison avec les protéines clientes médiées par ses co-chaperons2. Malgré un potentiel évident de Hsp90 en tant que cible thérapeutique, affiner sa fonction représente un grand défi. Il existe plusieurs inhibiteurs de Hsp90 ciblant la région de liaison de l’ATP N-terminal, qui ont été évalués dans des essais cliniques, mais aucun d’entre eux n’a été approuvé pour lacommercialisation 3. En raison de l’absence d’une poche de liaison au ligand4bien définie, le ciblage de la région C-terminale de Hsp90 a eu un succès limité4. Récemment, la perturbation des interactions Hsp90-cochaperone par de petites molécules a été étudiée comme stratégie alternative5. Le ciblage des interactions Hsp90-cochaperone ne provoquerait pas de réponse générale au stress cellulaire et offre la possibilité de réguler spécifiquement divers processus intracellulaires. Le pentapeptide MEEVD conservé à la terminaison C de Hsp90 est responsable de l’interaction avec le motif de répétition tétratricopeptide (TPR) des co-chaperons6. Sur les 736 protéines contenant des motifs TPR annotées dans la base de données des protéines humaines, environ 20 protéines différentes interagissent avec Hsp90 via ce peptide7. Les molécules en concurrence pour la liaison peptidique MEEVD perturberaient les interactions entre Hsp90 et les co-chaperons contenant un domaine TPR. Le site de liaison peptidique a une structure tertiaire similaire, mais l’homologie globale entre les différents domaines du motif TPR est relativement faible7,ce qui offre la possibilité d’identifier des molécules spécifiquement capables de bloquer les interactions entre Hsp90 et des co-chaperons particuliers à motif TPR. Parmi ces co-chaperons à motif TPR, la protéine de liaison FK506 (FKBP) 51 et FKBP52 sont des régulateurs de la signalisation des récepteurs hormonaux stéroïdiens (SHR) et impliqués dans plusieurs maladies hormono-dépendantes des stéroïdes, y compris le cancer, les maladies liées au stress, les maladies métaboliques et la maladie d’Alzheimer8. Bien que FKBP51 et FKBP52 partagent > similitude de séquence de 80%, leurs fonctions diffèrent: FKBP52 est un régulateur positif de l’activité SHR, tandis que FKBP51 est un régulateur négatif dans la plupart des cas8. Par conséquent, l’identification de molécules, bloquant spécifiquement les interactions entre Hsp90 et FKBP51 ou FKBP52, offre un potentiel thérapeutique prometteur pour les maladies connexes.
Unessai amplifié Luminescent Proximity Homogenous A(AlphaScreen) a été développé pour la première fois en 1994 par Ullman EF et al.9. Maintenant, il est largement utilisé pour détecter différents types d’interactions biologiques, telles que lepeptide 10,la protéine11,l’ADN12,l’ARN13et le sucre14. Dans cette technique, il existe deux types de perles (diamètre 200 nm), l’une est la perle donneuse et l’autre est la perle accepteuse. Les biomolécules sont immobilisées sur ces perles; leurs interactions biologiques rapprochent les perles donneuses et accepteuses. À 680 nm, un photosensibilisateur dans la perle du donneur éclaire et convertit l’oxygène en oxygène syté. Parce que l’oxygène syt a une courte durée de vie, il ne peut diffuser que jusqu’à 200 nm. Si la bille accepteur est à proximité, son dérivé de thioxène réagit avec l’oxygène singulet générant une chimiluminescence à 370 nm. Cette énergie active en outre les fluorophores dans la même bille accepteur pour émettre de la lumière à 520-620 nm15. Si les interactions biologiques sont perturbées, la perle accepteur et la perle donneuse ne peuvent pas atteindre la proximité, ce qui entraîne la désintégration de l’oxygène syulet et un faible signal produit.
Nous décrivons ici un protocole utilisant cette technique pour le criblage de petites molécules inhibant les interactions entre Hsp90 et les co-chaperons TPR, en particulier FKBP51 et FKBP52. Les 10 peptides longs d’acides aminés correspondant à Hsp90 extrême C-terminus sont attachés à des billes acceptrices. Les co-chaperons TPR purifiés marqués de la TPS interagissent avec les perles de donneurs liées au glutathion. Lorsque l’interaction entre les peptides dérivés de Hsp90 et les co-chaperons à motif TPR rassemble les billes, un signal amplifié est produit(Figure 1A). Si les petites molécules criblées peuvent inhiber les interactions entre Hsp90 et les co-chaperons à motif TPR, ce signal amplifié sera diminué(Figure 1B). Leur IC50 peut être calculé par mesure quantitative. Ce protocole peut être étendu à toutes les interactions chaperon -TPR-motif co-chaperon d’intérêt et est d’une grande importance dans le développement de nouvelles molécules, bloquant spécifiquement l’interaction entre Hsp90 et FKBP51 ou FKBP52.
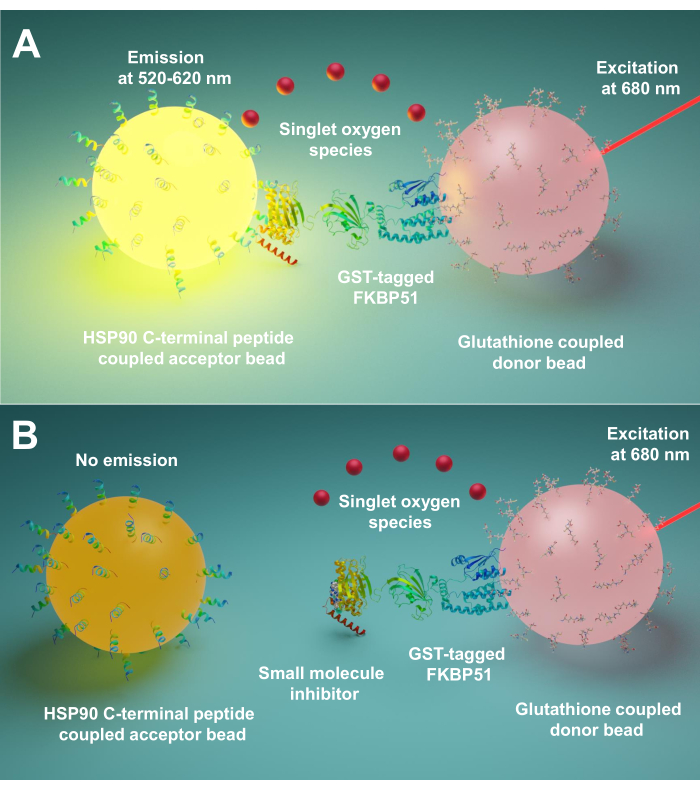
Figure 1: Principe de base de ce test. (A) La GST-FKBP51 purifiée interagit avec les billes de donneur liées au glutathion. Les 10 peptides longs d’acides aminés correspondant à l’extrême C-terminus de Hsp90 sont attachés à des billes acceptrices. L’interaction entre les peptides dérivés de Hsp90 et le domaine TPR de FKBP51 rapproche les perles donneuses et acceptrices. À 680 nm, un photosensibilisateur dans la perle du donneur éclaire et convertit l’oxygène en oxygène syté. Le dérivé de thioxène sur la bille accepteur réagit avec l’oxygène syt et génère une chimiluminescence à 370 nm. Cette énergie active en outre les fluorophores dans la même bille accepteur pour émettre de la lumière à 520-620 nm. (B) Lorsque de petites molécules inhibent les interactions entre Hsp90 et FKBP51, les perles donneuses et acceptatrices ne peuvent pas atteindre la proximité. Ensuite, l’oxygène syt avec une courte durée de vie se désintègre et aucun signal détectable n’est produit. Veuillez cliquer ici pour voir une version agrandie de cette figure.
