Den ikke-specifikke endocytiske vej for makropinocytose gør det muligt for celler at internalisere en række ekstracellulære komponenter, herunder næringsstoffer, proteiner, antigener og patogener, gennem bulkoptagelse af ekstracellulær væske og dets bestanddele1. Selvom det er vigtigt for biologien hos mange celletyper, beskrives makropinocytosevejen i stigende grad at spille en væsentlig rolle i tumorbiologi, hvor tumorceller gennem makropinocytisk optagelse er i stand til at overleve og sprede sig i nærvær af et næringsstofudtømt mikromiljø2,3. Optagelsen af ekstracellulære makromolekyler, herunder albumin og ekstracellulær matrix og nekrotisk celleaffald, giver en alternativ næringskilde til biomasseproduktion ved at skabe aminosyrer, sukkerarter, lipider og nukleotider gennem makropinosom og lysosomfusionsmedieret lastkatabolisme4,5,6,7,8.
Induktion og regulering af makropinocytose er kompleks og kan variere afhængigt af cellulær kontekst. Indtil videre er flere inducere af makropinocytose blevet identificeret og omfatter ligander, såsom epidermal vækstfaktor (EGF), blodpladeafledt vækstfaktor (PDGF), galectin-3 og Wnt3A9,10,11,12,13. Derudover kan dyrkningsbetingelser, der efterligner tumormikromiljøet, udløse aktivering af vejen. Pankreas duktal adenocarcinom (PDAC) tumorer er næringsstofberøvet, især for aminosyren glutamin, hvilket får både kræftceller og kræftassocierede fibroblaster (CAF’er) til at stole på makropinocytose for overlevelse7,13,14,15. Desuden kan tumorspændinger, såsom hypoxi og oxidativ stress, aktivere denne ådselsvej16. Ud over de mange ydre påvirkere, der kan inducere makropinocytose, styrer en række intracellulære veje makropinosomdannelse. Onkogen Ras-medieret transformation er tilstrækkelig til at starte det makropinocytiske maskineri, og flere kræfttyper udviser onkogen Ras-drevet konstitutiv makropinocytose4,5,9,17. Alternativt er vildtype Ras-aktivering og Ras-uafhængige veje blevet identificeret for at aktivere makropinocytose i kræftceller og CAF’er10,11,15,18. Anvendelsen af forskellige in vitro-modeller i kombination med inhibitorbehandlinger har resulteret i identifikation af flere makropinocytosemodulatorer, som omfatter natrium-hydrogenbyttere, den lille GTPase Rac1, phosphoinositid 3-kinase (PI3K), p21-aktiveret kinase (Pak) og AMP-aktiveret proteinkinase (AMPK)4,13,15 . I betragtning af de mange beskrevne faktorer og tilstande, der regulerer makropinocytose, er det imidlertid tænkeligt, at mange flere modulatorer og stimuli forbliver uopdagede. Identifikationen af nye modulatorer og stimuli kan lettes ved automatiseret vurdering af en lang række forhold i et enkelt eksperiment. Denne metode kan kaste lys over de faktorer, der er involveret i makropinosomdannelse og kan muliggøre opdagelsen af nye små molekyler eller biologiske lægemidler, der er målrettet mod denne vej.
Her har vi tilpasset vores tidligere etablerede protokol til bestemmelse af omfanget af makropinocytose i kræftceller in vitro til et 96-brønds mikropladeformat og automatiseret billeddannelse og kvantificering19,20. Denne protokol er baseret på fluorescerende mikroskopi, som er blevet en standard på området til bestemmelse af makropinocytose in vitro og in vivo4,5,6,7,9,10,11,12,13,15,16,17,18, 19,20,21,22. Makropinosomer kan skelnes fra andre endocytiske veje gennem deres evne til at internalisere store makromolekyler, såsom højmolekylær dextran (dvs. 70 kDa)2,3,4,20,21,22,23. Således kan makropinosomer defineres ved optagelse af ekstracellulært administreret fluorofor-mærket 70 kDa dextran. Som et resultat manifesterer makropinocytiske vesikler sig som intracellulære klynger af fluorescerende puncta med størrelser fra 0,2-5 μm. Disse puncta kan mikroskopisk afbildes og efterfølgende kvantificeres for at bestemme omfanget af makropinocytose i cellen – ‘det makropinocytiske indeks’.
I denne protokol beskrives de væsentlige trin til visualisering af makropinosomer i klæbende celler in vitro på en 96-brønds mikroplade og dæksedler ved hjælp af standard laboratorieudstyr (figur 1). Derudover gives anvisningerne til automatisering af billedindsamling og kvantificering af det makropinocytiske indeks ved hjælp af en cellebilleddannelsespladelæser med flere tilstande. Denne automatisering reducerer tid, omkostninger og kræfter sammenlignet med vores tidligere beskrevne protokoller19,20. Derudover undgår det utilsigtet forudindtaget billedindsamling og analyse og forbedrer derved reproducerbarheden og pålideligheden. Denne metode kan let tilpasses forskellige celletyper eller pladelæsere eller bruges til at bestemme alternative makropinosomfunktioner, såsom størrelse, antal og placering. Den heri beskrevne metode er især velegnet til screening af cellekulturbetingelser, der inducerer makropinocytose, identifikation af nye modulatorer eller optimering af lægemiddelkoncentrationer af kendte hæmmere.
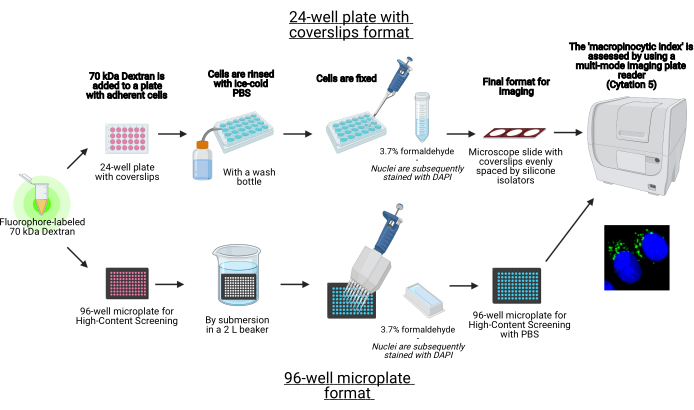
Figur 1: Skematisk over det automatiserede assay til bestemmelse af det ‘makropinocytiske indeks’ i klæbende celler. Oprettet ved hjælp af BioRender. Klik her for at se en større version af denne figur.
