Using this protocol, the cell number and the concentrations of various OxPHOS-targeting drugs (used in the extracellular flux assays) were optimized to measure ECAR and OCR of HSPCs isolated from 24-week-old female C57BL/6 mice. First, the glycolysis stress test was performed to optimize cell number and oligomycin concentration. A varying number of HSPCs per well ranging from 5 × 104 to 2.5 × 105 were used in this assay. As shown in Figure 2A and Figure 2C, the non-glycolytic acidification rate rises with increased cell number from 5 × 104 to 2.5 × 105 but increases only minimally between 2 × 105 to 2.5 × 105 cells. As expected, injection of 10 mM glucose stimulates glycolysis at all cell numbers, with a maximum increase in ECAR observed at 2.5 × 105 cells per well. However, injection of 2 μM of oligomycin does not further increase ECAR, as otherwise expected. The use of 1 μM oligomycin yielded similar results (not shown). These results could be interpreted to indicate that these oligomycin concentrations were not optimal in these assays.
However, OCR data obtained in the same set of glycolysis stress tests show that both tested oligomycin concentrations (1 μM and 2 μM) were indeed effective, as suggested by the significant drop in OCR following oligomycin injection due to complex V inhibition (see Figure 2B for 2 μM oligomycin; not shown for 1 μM oligomycin). At both tested oligomycin concentrations, maximal decreases in OCR were observed at 2.5 × 105 cells per well. Finally, injection of 50 mM 2-DG resulted in a significant reduction of ECAR, implying that glycolysis is the source of ECAR observed in these experiments. Taken together, it may be concluded that HSPCs achieve maximum glycolysis following glucose injection in these assays, and they possess little-to-no glycolytic reserve. This indicates that, like purified HSCs, lineage-negative HSPCs, which include the HSC fraction used in this assay, also rely predominantly on glycolysis for their ATP production. In cells with a high glycolytic rate,there may not be a significant increase in ATP demand upon oligomycin-mediated inhibition of mitochondrial ATP production. Cells could easily manage the loss of mitochondrial ATP without further upregulating glycolysis19. Glycolysis stress test parameters-non-glycolytic acidification, glycolysis, glycolytic capacity, and glycolytic reserve as shown in Figure 2C-were calculated as described previously in the introduction and protocol sections. Based on these data, 2.5 × 105 cells per well and 2 μM of oligomycin were selected for further studies.
The mitochondrial stress test was used for the optimization of FCCP concentration. In this assay, 2.5 × 105 HSPCs were used per well, as optimized previously via the glycolysis stress test. As shown in Figure 3A, the assay begins with the measurement of baseline OCR followed by 2 μM oligomycin injection, causing a significant reduction in OCR via the inhibition of complex V. Following post-oligomycin injection OCR measurements, FCCP was injected at varying concentrations: 0.5 μM, 1 μM, 1.5 μM, and 2 μM. As indicated earlier, FCCP reverses the oligomycin-induced repression of electron flow through ETC by uncoupling proton transport from OxPHOS and forces complex IV to consume oxygen maximally. As shown in Figure 3A, FCCP stimulates OCR in HSPCs in a dose-dependent manner with a maximal increase in OCR observed at 2 μM of FCCP. Finally, a mixture of 0.5 μM rotenone and 0.5 μM antimycin A was injected, which completely shuts down electron flow through the ETC, and OCR decreases to its minimal level. OCR measured after rotenone and antimycin A injection corresponds to non-mitochondrial oxygen consumption. Other mitochondrial stress test parameters-basal respiration, maximal respiration, proton leak, ATP production, spare respiratory capacity, and coupling efficiency as shown in Figure 3B-were calculated as described previously in the introduction and protocol sections. Finally, 2 μM of FCCP was selected for further studies.
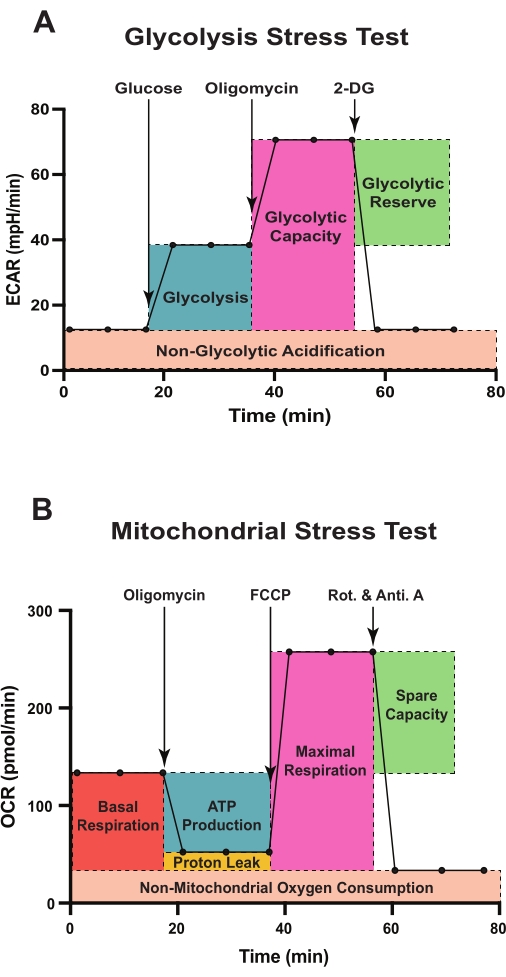
Figure 1: Schematic representation of assessment of glycolytic function and mitochondrial respiration using the extracellular flux analyzer. Sequences and injection timings of various effector compounds used in the glycolytic stress test (A) and the mitochondrial stress test (B) are shown. (A) Glycolytic parameters, glycolysis, glycolytic capacity, glycolytic reserve, and non-glycolytic acidification, and (B) mitochondrial function parameters, basal respiration, ATP-linked respiration, maximal respiration, non-mitochondrial oxygen consumption, proton leak, and spare respiratory capacity are outlined. Abbreviations: ECAR = extracellular acidification rate; 2-DG = 2-deoxy-D-glucose; OCR = oxygen consumption rate; FCCP = carbonyl cyanide-4 (trifluoromethoxy) phenylhydrazone; Rot. = rotenone; Anti. A = antimycin A. Please click here to view a larger version of this figure.
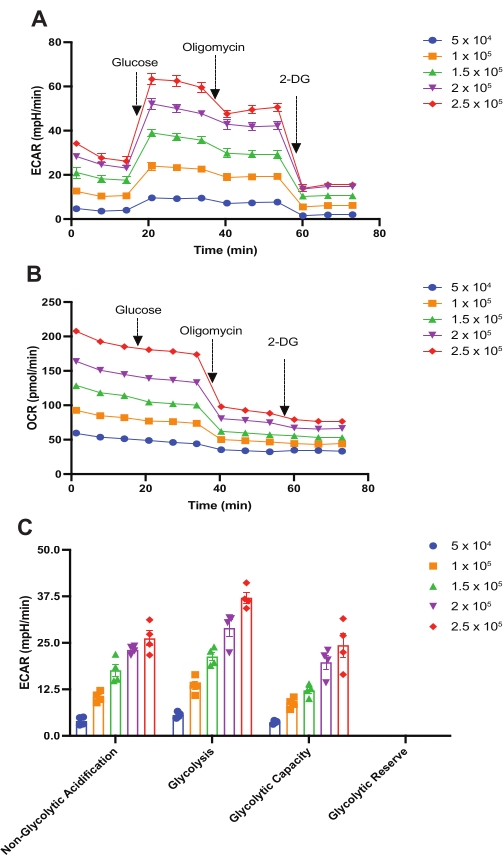
Figure 2: Assessment of the glycolytic function in mouse HSPCs. The glycolysis stress test was performed to measure (A) the extracellular acidification rates (ECAR, mpH/min) and (B) the oxygen consumption rates (OCR, pmol/min) of HSPCs isolated from 24-week-old female C57BL/6 mice. At times indicated, glucose (10 mM), oligomycin (2 µM), and 2-DG (50 mM) were injected. (C) ECAR is calculated as non-glycolytic acidification, glycolysis, glycolytic capacity, and glycolytic reserve per 5 × 104, 1 × 105, 1.5 × 105, 2 × 105, and 2.5 × 105 HSPCs/well. Data are presented as mean ± SEM, n = 4. Abbreviations: HSPCs = hematopoietic stem and primitive progenitor cells; ECAR = extracellular acidification rate; 2-DG = 2-deoxy-D-glucose; OCR = oxygen consumption rate. Please click here to view a larger version of this figure.
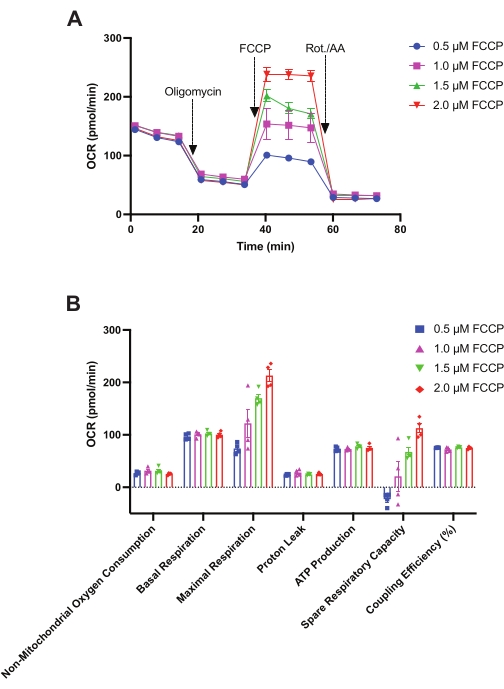
Figure 3: Assessment of the mitochondrial respiration in mouse HSPCs. A mitochondrial stress test was performed to measure (A) the oxygen consumption rates (pmol/min) of HSPCs isolated from 24-week-old female C57BL/6 mice. At times indicated, oligomycin (2 µM), FCCP (0.5 µM, 1 µM, 1.5 µM, 2 µM), and rotenone and antimycin A (Rot./AA, 0.5 µM each) were injected. (B) OCR is calculated as non-mitochondrial oxygen consumption, basal respiration, proton leak, maximal respiration (per 0.5 µM, 1 µM, 1.5 µM, and 2 µM FCCP), ATP production, spare respiratory capacity (per 0.5 µM, 1 µM, 1.5 µM, and 2 µM FCCP), and coupling efficiency per 2.5 × 105 HSPCs/well. Data are presented as mean ± SEM, n = 4. Abbreviations: HSPCs = hematopoietic stem and primitive progenitor cells; OCR = oxygen consumption rate; FCCP = carbonyl cyanide-4 (trifluoromethoxy) phenylhydrazone; Rot. = rotenone; AA = antimycin A. Please click here to view a larger version of this figure.
| Injection Port | Port volume | Injecting compound (10x concentrated) | Final compound concentration in the well |
| A | 20 μL | Glucose (100 mM) | 10 mM |
| B | 22 μL | Oligomycin (20 μM) | 2 μM |
| C | 25 μL | 2-DG (500 mM) | 50 mM |
Table 1: Injection strategy for the glycolysis stress test. Abbreviation: 2-DG = 2-deoxy-D-glucose.
| 1 | Calibration | ||||
| 2 | Equilibration | ||||
| 3 | Injections and measurements | ||||
| Cycles | Mix | Wait | Measure | ||
| Baseline (Non-glycolytic acidification) | 3 times | 3 min | 0 min | 3 min | |
| Inject Port A: Glucose | 3 times | 3 min | 0 min | 3 min | |
| Inject Port B: Oligomycin | 3 times | 3 min | 0 min | 3 min | |
| Inject Port C: 2-DG | 3 times | 3 min | 0 min | 3 min | |
Table 2: Extracellular flux analyzer program for the glycolysis stress test. Abbreviation: 2-DG = 2-deoxy-D-glucose.
| Injection Port | Port volume | Injecting compound (10x concentrated) | Final compound concentration in the well |
| A | 20 μL | Oligomycin (20 μM) | 2 μM |
| B | 22 μL | FCCP (20 μM) | 2 μM |
| C | 25 μL | Rotenone (5 μM) + antimycin A (5 μM) | 0.5 μM each |
Table 3: Injection strategy for the mitochondrial stress test. Abbreviation: FCCP = carbonyl cyanide-4 (trifluoromethoxy) phenylhydrazone.
| 1 | Calibration | ||||
| 2 | Equilibration | ||||
| 3 | Injections and measurements | ||||
| Cycles | Mix | Wait | Measure | ||
| Baseline measurements | 3 times | 3 min | 0 min | 3 min | |
| Inject Port A: Oligomycin | 3 times | 3 min | 0 min | 3 min | |
| Inject Port B: FCCP | 3 times | 3 min | 0 min | 3 min | |
| Inject Port C: Rotenone & Antimycin A | 3 times | 3 min | 0 min | 3 min | |
Table 4: Extracellular flux analyzer program for the mitochondrial stress test. Abbreviation: FCCP = carbonyl cyanide-4 (trifluoromethoxy) phenylhydrazone.
