Målinger av Cilia Beat Frequency (CBF) og mønster er etablert som diagnostiske verktøy for luftveissykdommer som primær ciliær dyskinesi (PCD)1. Ved cystisk fibrose (CF) forårsaker dysfunksjon av CF Transmembrane conductance Regulator (CFTR)-kloridkanalen dehydrering av væsken på luftveisoverflaten og nedsatt slimclearance2. Ciliærfunksjonen er undersøkt in vitro i primære luftveiscellemodeller som en indikator på CFTR-kanalaktivitet3. Imidlertid eksisterer det betydelig pasient-til-pasient-variasjon i CBF som respons på CFTR-modulerende legemidler, selv for pasienter med de samme CFTR-mutasjonene 3. Videre er virkningen av dysfunksjonell CFTR-regulert kloridsekresjon på ciliær funksjon dårlig forstått. Det er for tiden ingen omfattende protokoll som demonstrerer prøvepreparering av in vitro luftveismodeller, bildeinnsamling og analyse av CBF.
Neseepitelark isolert fra neseslimhinnebørsting brukes direkte til målinger av ciliær funksjon for PCD-diagnose4. Likevel, mens det ikke er kontroll over størrelsen eller kvaliteten på neseepitelarkene som er oppnådd, varierer CBF avhengig av om det måles på enkeltceller eller celleark og på epitelark cilierte kanter som er forstyrret eller uforstyrret5. Som sådan kan sekundære dyskinesier forårsaket av skade på celler under samlingen av neseslimhinnebørster påvirke CBF. Primær cellekultur av neseepitelceller og deres differensiering ved luft-væskegrensesnitt (ALI) eller i tredimensjonal kjellermembranmatrise i cilierte luftveisepitelorganoider gir opphav til cilia som er fri for sekundære dyskinesier 4,6,7,8. Luftveisepitelceller differensiert ved ALI (heretter kalt ALI-modeller) har blitt ansett som et viktig sekundært diagnostisk hjelpemiddel som replikerer ciliary beat-mønstrene og frekvensen av ex vivo nasal mucosal brushings6 og muliggjør analyse av ciliær ultrastruktur, beatmønster og beatfrekvens samtidig som pasientspesifikke defekter opprettholdes9 . Likevel eksisterer det uoverensstemmelser i metodene som brukes til å lage disse pseudostratifiserte, mucociliary differensierte cellemodellene. Ulike kulturutvidelses- eller differensieringsprotokoller kan indusere distinkte epitelfenotyper (ciliert eller sekretorisk)10 og resultere i signifikante forskjeller i CBF11. CBF har blitt kvantifisert i neseepitelbørster 4,6,12,13,14,15,16, luftveisepitelorganoider 14,17,18 og ALI-modeller 3,4,6,13,19,20, 21. Likevel, blant disse protokollene, er det store variasjoner, og ofte er mange parametere ikke kontrollert for. For eksempel, i noen studier, er CBF avbildet in situ mens cellene i ALI-modellen forblir på den permeable støtteinnsatsen 3,19,20,21, mens andre skraper cellene fra den permeable støtteinnsatsen og avbilder dem suspendert i media 4,6,13.
Videre er den bredere anvendelsen av teknikker som måler ciliær funksjon begrenset av den ekstreme følsomheten til ciliær funksjon for endringer i miljøfaktorer. Miljøfaktorer som temperatur22, fuktighet 23,24 og pH 25,26 påvirker ciliærfunksjonen og må reguleres for å kvantifisere CBF nøyaktig. De ulike fysiologiske parametrene som brukes på tvers av ulike laboratorier og hvordan de påvirker CBF har blitt gjennomgått tidligere27.
Ulike bildeteknologier og tilnærminger til CBF-målinger er rapportert i litteraturen. For PCD-diagnostikk brukes videomikroskopi til å måle ciliær funksjon28,29. Nylig ble en videoanalysealgoritme basert på differensiell dynamisk mikroskopi brukt til å kvantifisere både CBF og cilia-koordinasjon i luftveisepitelcelle ALI-modeller 3,30. Denne metoden gjør det mulig å karakterisere ciliær juling i luftveisepitelceller på en rask og helautomatisk måte, uten behov for å segmentere eller velge regioner. Ulike metoder for avbildning og kvantifisering av CBF kan bidra til forskjeller rapportert i CBF i litteraturen (tilleggsfil 1).
En protokoll fra kultur til kvantifisering for å effektivisere eksisterende metoder, standardisering av kulturforhold og bildeoppkjøp, utført under strenge miljøkontrollerte forhold, vil muliggjøre konsistent, reproduserbar kvantifisering av CBF innenfor og mellom individer.
Denne protokollen gir en fullstendig beskrivelse av samlingen av epitelceller, ekspansjons- og differensieringskulturforhold og kvantifisering av CBF i tre forskjellige luftveisepitelcellemodellsystemer av nasal opprinnelse: 1) innfødte epitelark, 2) ALI-modeller avbildet på permeable støtteinnlegg og 3) Ekstracellulær matrise (ECM)-innebygde tredimensjonale organoider (figur 1 ). Neseepitelceller oppnådd fra nasal dårligere turbinatbørster brukes som representanter for luftveisepitelet siden de er et effektivt surrogat for bronkialepitelceller31 mens de overvinner den invasive prosedyren forbundet med å samle bronkial børster. Metoden Conditional Reprogramming Cell (CRC) brukes til å utvide primære luftveisepitelceller for opprettelse av ALI-modeller og tredimensjonale organoider. Betinget omprogrammering av luftveisepitelceller til stamcellelignende tilstand induseres av samkultur med vekstarrestert fibroblastmatercellesystem og Rho-assosiert kinase (ROCK) hemmer32. Det er viktig at CRC-metoden øker populasjonsdoblingen i luftveisepitelceller samtidig som de beholder sitt vevsspesifikke differensieringspotensial33,34. I alle luftveisepitelcellemodeller fanges ciliary-funksjonen i et temperaturkontrollert kammer ved hjelp av et høyhastighets videokamera med standardiserte bildeinnsamlingsinnstillinger. Spesialbygde skript brukes til kvantifisering av CBF.
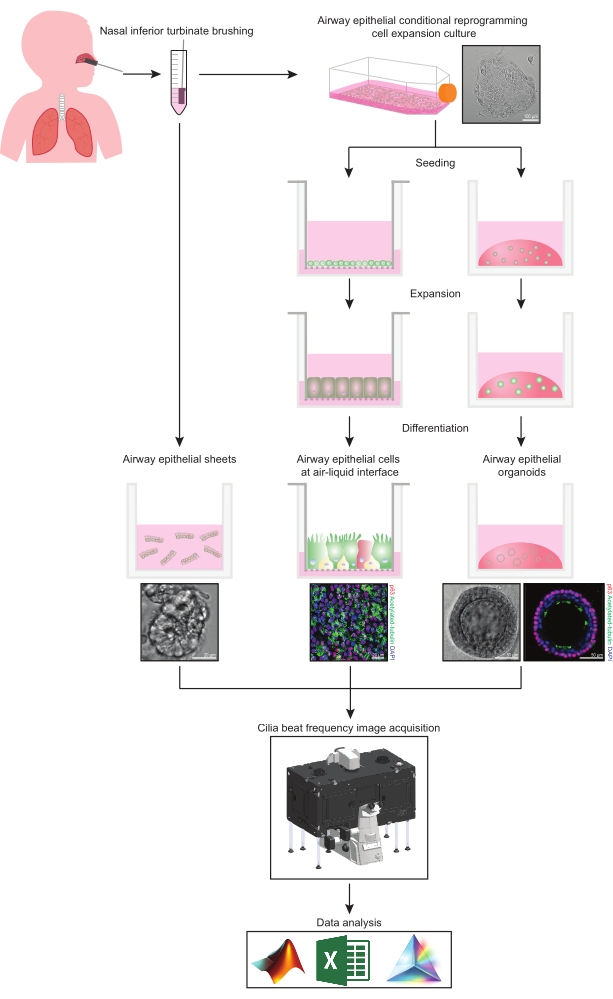
Figur 1: Skjematisk arbeidsflyt. Etter børsting av deltakernes nasale nedre turbinat, brukes luftveisepitelceller på en av to måter. Enten isoleres luftveisepitelark, og cilia beat-frekvensen avbildes umiddelbart, eller luftveienes epitelceller utvides via den betingede omprogrammeringscellemetoden. CRC-ekspanderte luftveisepitelceller differensieres for å etablere luftveisepitelceller ved et luft-væskegrensesnitt eller luftveisepitelorganoidkulturer. Avbildning av ciliær beatfrekvens oppnås ved hjelp av et levende celleavbildningsmikroskop med et varme- og fuktighetsmiljøkammer og et raskt bildefrekvens (>100Hz) vitenskapelig kamera. Dataanalyse utføres ved hjelp av spesialbygde skript. Klikk her for å se en større versjon av dette tallet.
