ويرافق تكوين العظام بواسطة الأرومة العظمية تدمير منسق أو ارتشاف العظام بواسطة الخلايا الآكلة للعظام. التوازن بين تكوين العظام العظمية الأرومية وارتشاف الخلايا العظمية هو عملية مقترنة تصف دوران العظام أو إعادة تشكيلها ، وهو أمر ضروري لتوازن الهيكل العظمي. يؤدي خلل العظام إلى ضعف تكوين العظام ويؤدي إلى أمراض مختلفة ، بما في ذلك هشاشة العظام1،2،3. يؤدي التمايز خارج الجسم الحي / في المختبر للخلايا الجذعية اللحمية لنخاع العظم (BMSCs) إلى سلائف الخلايا العظمية العظمية والأرومات العظمية الناضجة إلى تكوين وترسب مصفوفة العظام المعدنية في وعاء الاستزراع بمرور الوقت4،5،6. يتطلب تكوين العظام هذا بواسطة الأرومة العظمية كمية كبيرة من الطاقة الخلوية. على وجه التحديد ، ثبت أن تخليق الكولاجين وإفرازه يعتمدان بشكل كبير على ATP الخلوي: نسب ADP ، ومن المفترض أن يتطلب الاتجار بالحويصلات المعدنية وإفرازهاATP إضافيا 7,8,9,10,11. وقد أثبت العديد من الباحثين أن عملية تكوين الخلايا الأرومية العظمية ووظيفة الأرومة العظمية تتطلب إمدادات كافية من الطاقة لتلبية الطلب الأيضي لتشكيل العظام12،13،14،15،16. لذلك ، فإن الهدف من هذه الطريقة هو توصيف حالة الطاقة الحيوية للخلايا اللحمية الأولية الفئرانية طوال تمايز الخلايا العظمية باستخدام محلل التدفق الأيضي للخلية في الوقت الفعلي. تساعد هذه التقنيات في تطوير فهم أفضل لتوازن الهيكل العظمي ، مما قد يؤدي في النهاية إلى تطوير خيارات علاجية جديدة قادرة على تحسين اضطرابات الهيكل العظمي.
يمكن استخدام محلل التدفق الأيضي للخلايا في الوقت الفعلي لقياس معدل استهلاك الأكسجين (OCR) ومعدل التحمض خارج الخلية (ECAR) للخلايا العظمية الحية ، والتي تتوافق مع إنتاج الميتوكوندريا و ATP المحللة للسكر ، على التوالي. من الأمور الأساسية لهذه المنهجية حقيقة أن أيون H+ واحد لكل لاكتات يتم إطلاقه أثناء تحلل السكر في تحويل الجلوكوز إلى لاكتات ، مما يغير درجة الحموضة في الوسائط المنعكسة في قيم ECAR. على العكس من ذلك ، خلال دورة TCA (حمض ثلاثي الكربوكسيليك) ، تنتج الفسفرة التأكسدية عبر الميتوكوندريا ثاني أكسيد الكربون2 عن طريق استخدام أو استهلاك الأكسجين ، وبالتالي فإن مراقبة OCR تعكس عملية التمثيل الغذائي هذه. يقيس المحلل كلا من OCR و ECAR في البيئة الدقيقة خارج الخلية في وقت واحد وفي الوقت الفعلي ، مما يسمح بإمكانات هائلة عند دراسة الطاقة الحيوية الخلوية 6,17. بالإضافة إلى ذلك ، فإن إجراء هذه الفحوصات بسيط نسبيا ويمكن تخصيصه بسهولة اعتمادا على الهدف التجريبي. تم استخدام تقنيات مماثلة لزيادة فهم التنظيم الأيضي للخلايا التائية للجهاز المناعي18,19 ، وبدء السرطان ، والتقدم 20 ، إلى جانب أنواع متعددة من الخلايا الأخرى التي تساهم في متلازمات التمثيل الغذائي21,22.
تشمل مزايا محلل التدفق الأيضي في الوقت الفعلي على التقنيات البديلة (1) القدرة على قياس الطاقة الحيوية الخلوية للخلايا الحية في الوقت الفعلي ، (2) القدرة على إجراء الفحص مع عدد صغير نسبيا من الخلايا (يتطلب ما يصل إلى 5000 خلية) ، (3) منافذ الحقن لمعالجة العلاجات المتعددة بالتوازي في نظام 96 بئرا عالي الإنتاجية ، (4) استخدام جهاز تصوير الخلايا الآلي الخالي من الملصقات المشعة للتطبيع18 ، 23,24. تهدف الطرق التالية إلى توفير وصف معمم ولكن مفصل لمراقبة الطاقة الحيوية الخلوية في BMSCs الفئران طوال تمايز الخلايا العظمية باستخدام المحلل. وسوف تشمل الفحوص التي تجرى بشكل روتيني. ومع ذلك ، كما هو الحال مع العديد من التقنيات والأساليب ، من المشجع للغاية أن تحدد المختبرات الفردية تفاصيل محددة لتجاربها.
اختيار المقايسة وأنواع مختلفة من المقايسات المتاحة: تتوفر مجموعة واسعة من مجموعات المقايسة والكواشف لدراسة الطاقة الحيوية للخلايا مع ضمان موثوقية واتساق النتائج التجريبية. بالإضافة إلى ذلك ، يوفر برنامج سطح المكتب أيضا قوالب فحص يمكن تخصيصها بسهولة. يمكن تعريف الفحص بناء على احتياجات المستخدم لقياس معلمات التمثيل الغذائي المختلفة. يمكن تعديل هذه الفحوصات بطرق مختلفة بناء على الهدف التجريبي و / أو السؤال العلمي. على سبيل المثال ، مع أربعة منافذ حقن ، يمكن حقن مركبات متعددة في وسائط الفحص لتحليل الاستجابة الخلوية الخاصة بكل مسار استقلابي.
اختبار النمط الظاهري لطاقة الخلية: يقيس هذا الفحص النمط الظاهري الأيضي للخلايا الحية وإمكانات التمثيل الغذائي. يوصى أيضا بإجراء هذا الفحص كخطوة أولى للحصول على فكرة عامة عن التمثيل الغذائي الخاص بالمسار. يتم حقن خليط من أوليغوميسين A-مثبط من سينثاز ATP وسيانيد الكربونيل 4-(ثلاثي فلوروميثوكسي) فينيل هيدرازون (FCCP) – وهو عامل فك ارتباط الميتوكوندريا لفهم إمكانات طاقة الخلية. حقن oligomycin A يمنع تخليق ATP ، مما يؤدي إلى زيادة في معدل تحلل السكر (ECAR) لتمكين الخلايا من تلبية احتياجاتها من الطاقة ؛ من ناحية أخرى ، يؤدي حقن FCCP إلى ارتفاع OCR بسبب إزالة استقطاب غشاء الميتوكوندريا. في الأساس ، يصور هذا الفحص التنفس الأيضي الأساسي ، وبعد الحقن المزدوج أو الدفع أو الإجهاد ، الاستجابة الأيضية. بناء على هذه المعلمات ، يقوم البرنامج بعد ذلك برسم OCR و ECAR للخلايا عن طريق تصنيف الخلايا على أنها حالة هوائية أو هادئة أو تحلل السكر أو نشطة بمرور الوقت25,26.
ATP في الوقت الحقيقي معدل الإنتاج: هذا يقيس إنتاج ATP الخلوي في وقت واحد من تحلل السكر والتنفس الميتوكوندريا. يقيس هذا الفحص كميا التحولات الأيضية من مساري الطاقة ويوفر بيانات عن معدلات إنتاج ATP للميتوكوندريا وتحلل السكر بمرور الوقت. يحصل الفحص على بيانات OCR و ECAR القاعدية متبوعة بحساب معدل إنتاج ATP الميتوكوندريا من خلال حقن oligomycin A ومعدل إنتاج ATP المحلل للسكر من خلال حقن خليط rotenone + antimycin A (التثبيط الكلي لوظيفة الميتوكوندريا) ، مما يؤدي إلى تحمض الميتوكوندريا17,27.
اختبار إجهاد الميتوكوندريا الخلوية (أو اختبار إجهاد ميتو الخلوي): هذا يقيس وظيفة الميتوكوندريا من خلال التنفس المرتبط ب ATP ، ويحدد كمية الطاقة الحيوية الخلوية ، ويحدد خلل الميتوكوندريا ، ويقيس استجابة الخلايا للإجهاد. يمكن الحصول على معلمات مختلفة ، بما في ذلك القدرة التنفسية القاعدية والاحتياطية ، والتنفس المرتبط ب ATP ، والتنفس الأقصى ، واستهلاك الأكسجين غير الميتوكوندريا ، في فحص واحد. يتضمن هذا الفحص حقن متسلسلة من oligomycin A ، FCCP (عامل فك ارتباط الميتوكوندريا) ، وهو مزيج من مثبطات الروتينون / antimycin A لتحليل تأثير هذه بكفاءة على وظيفة الميتوكوندريا28.
اختبار مرونة ميتو للوقود المرن: هذا يقيس معدل التنفس الميتوكوندريا عن طريق أكسدة وقود الميتوكوندريا الرئيسي الثلاثة من خلال وجود وغياب مثبطاتها. يساعد التثبيط المتسلسل للجلوكوز والجلوتامين والأحماض الدهنية في قياس اعتماد الخلايا وقدرتها ومرونتها واعتماد الخلايا في مسارات خلوية مختلفة لتلبية الطلب على الطاقة. عندما لا تستطيع الميتوكوندريا تلبية متطلبات المسار المسدود للاهتمام عن طريق أكسدة أنواع الوقود الأخرى ، تدخل الخلايا في حالة تبعية. يتم حساب قدرة الخلايا عن طريق تثبيط المسارين البديلين الآخرين متبوعا بتثبيط مسار الاهتمام. تساعد مرونة الخلايا في فهم قدرة الميتوكوندريا على تعويض وتلبية احتياجات الوقود للمسار المثبط. يتم حسابه عن طريق طرح تبعية الخلايا من قدرة الخلايا. يتم استخدام ثلاثة مثبطات مختلفة بشكل مستقل أو كمزيج من اثنين لحساب معلمات الفحص بشكل فعال. 2-سيانو-3-(1-فينيل-1H-إندول-3-يل)-2-حمض البروبينويك (UK5099) يمنع أكسدة الجلوكوز عن طريق منع الناقل البيروفات في تحلل السكر. Bis-2-(5-phenylacetamido-1,3,4-thiadiazol-2-yl) (BPTES) يمنع كبريتيد الإيثيل مسار أكسدة الجلوتامين ، ويمنع etomoxir أكسدة الأحماض الدهنية طويلة السلسلة29.
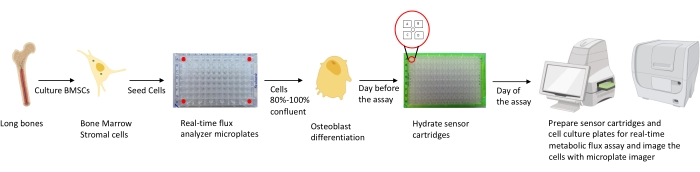
الشكل 1: التمثيل التخطيطي لمنهجية زراعة وإعداد الخلايا العظمية للتحليل. يتم عزل المورين BMSCs من العظام الطويلة ، وزرعها ، وزرعها في صفائح 96 بئرا بكثافة 25000 خلية / بئر. يبدأ زراعة هذه الخلايا في وسائط Osteoblast المحددة عندما تصل إلى 80٪ -100٪ من الالتقاء لبدء تمايزها. يتم إجراء الفحوصات في مراحل مختلفة من التمايز. يتم ترطيب ألواح الخرطوشة قبل يوم واحد من الفحص. في يوم الفحص ، يتم حقن مثبطات مختلفة في منافذ خراطيش المستشعر بناء على متطلبات الفحص ، ويضاف مخزن مؤقت للمعايرة إلى لوحة المعايرة المكونة من 96 بئرا. بعد المعايرة ، يتم إجراء اختبار التدفق الأيضي للخلية في الوقت الفعلي ، يليه تصوير الصفيحة الدقيقة لزراعة الخلايا باستخدام جهاز تصوير الصفائح الدقيقة لتطبيع بيانات محلل التدفق الأيضي للخلية في الوقت الفعلي مع عدد الخلايا. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
