Kaspazlar, başlatıcı kaspazlar ve cellat kaspazlar olarak gruplandırılabilen bir sistein aspartat proteazlar ailesidir. Cellater kaspazlar kaspaz-3, -6 ve -7’den oluşur. Doğal olarak hücrelerde dimerler olarak bulunurlar ve apoptoz1’i yürütmek için başlatıcı kaspazlar tarafından bölünürler. Başlatıcı kaspazlar insan kaspazı-1, -2, -4, -5, -8, -9, -10 ve -12’yi içerir. Yakınlığa bağlı dimerizasyon ile aktive olan ve otoproteolitik bölünme 2,3 ile stabilize edilen inaktif zimojenler (pro-kaspazlar) olarak bulunurlar. İnflamatuar kaspazlar, başlatıcı kaspaz2’nin bir alt kümesidir ve insanlarda kaspaz-1, -4, -5 ve -12’yi ve fare 4,5’te kaspaz-1, -11 ve -12’yi kapsar. Apoptotik bir rolden ziyade, inflamasyonda merkezi bir rol oynarlar. Patojenik istilacılara yanıt olarak salınan ilk sitokinler olan pro-interlökin (IL)-1β ve pro-IL-18 6,7’nin proteolitik işlenmesine ve salgılanmasına aracılık ederler 8,9. Caspase-1, aktivasyon platformuna işe alındıktan sonra etkinleştirilir; inflamatuar olarak adlandırılan büyük bir moleküler ağırlıklı protein kompleksi (Şekil 1A)10. Kaspaz-4, -5 ve -11’in dimerizasyonu, kanonik olmayan bir enflamatuar yol11,12 aracılığıyla bu platformlardan bağımsız olarak gerçekleşir.
Kanonik enflamatuarlar, bir enflamatuar sensör proteini, adaptör protein ASC (CARD içeren apoptozla ilişkili leke benzeri protein) ve efektör protein kaspaz-110’dan oluşan sitozolik multimerik protein kompleksleridir. En iyi çalışılmış kanonik enflamatuarlar, bir pirin alanı (NLRP), NLRP1 ve NLRP3 içeren NOD benzeri reseptör ailesi, bir CARD (NLRC), NLRC4 içeren NLR ailesi ve melanom 2’de (AIM2) bulunmayanlardır. Her biri bir pirin etki alanı, bir CARD veya her iki etki alanı içerir. CARD etki alanı, CARD içeren kaspazlar ve bunların yukarı akış aktivatörleri arasındaki etkileşime aracılık eder. Bu nedenle, bir N-terminal pirin alanı (PYD) ve bir C-terminal CARD motifi 13,14’ten oluşan iskele molekülü ASC, NLRP1 10, NLRP315 ve AIM216 enflamatuarlarına kaspaz-1’in alınması için gereklidir.
Her enflamatuar, farklı pro-inflamatuar uyaranları tanıyan benzersiz sensör proteininden sonra adlandırılır (Şekil 1B). Bu yolun aktivatörlerine kanonik uyaranlar denir. Enflamatuarlar, mikrobiyal bileşenler ve doku stresi için sensörler görevi görür ve enflamatuar kaspazların aktivasyonu yoluyla sağlam bir enflamatuar yanıtı tetiklemek için bir araya gelir17. Enflamatuar düzenek, ana substratları pro-IL-1β ve pro-IL-18’in olgunlaşmasına ve salgılanmasına aracılık etmek için kaspaz-1 aktivasyonunu başlatır. Bu işlem iki adımlı bir mekanizma ile gerçekleşir. İlk olarak, bir astarlama uyaranı, NF-κB yolunun aktivasyonu yoluyla bazı enflamatuar proteinlerin ve pro-IL-1β’nin ekspresyonunu düzenler. İkincisi, hücre içi (kanonik) bir uyaran, enflamatuar montaja ve prokaspaz-1 6,7’nin işe alınmasına neden olur.
Kaspaz-4 ve kaspaz-5, murin kaspaz-11 11’in insan ortologlarıdır. Gram-negatif bakterilerin dış zarında bulunan bir molekül olan hücre içi lipopolisakkarit (LPS) 18,19,20 ve kırmızı kan hücresi hemolizi 21’in bir ürünü olan hücre dışı heme ile enflamatuardan bağımsız bir şekilde aktive edilirler. LPS’nin doğrudan bu proteinlerin CARD motifine bağlandığı ve oligomerizasyonlarını indüklediği öne sürülmüştür20. Kaspaz-4 veya kaspaz-5’in aktivasyonu, gözenek oluşturan protein gasdermin D’nin (GSDMD) bölünmesi yoluyla piroptoz adı verilen enflamatuar bir hücre ölümü formunu indükleyerek IL-1β salınımını teşvik eder18,19. Ek olarak, kaspaz-4 ve GSDMD aracılı piroptotik ölümden kaynaklanan potasyum iyonlarının efflüksü, NLRP3 enflamatuarının aktivasyonunu ve ardından kaspaz-122,23’ün aktivasyonunu indükler. Bu nedenle, kaspaz-4, -5 ve -11, spesifik uyaranlar 11,24’e yanıt olarak piroptoz ve kaspaz-1 aktivasyonunu indükleyebilen LPS için hücre içi sensörler olarak kabul edilir.
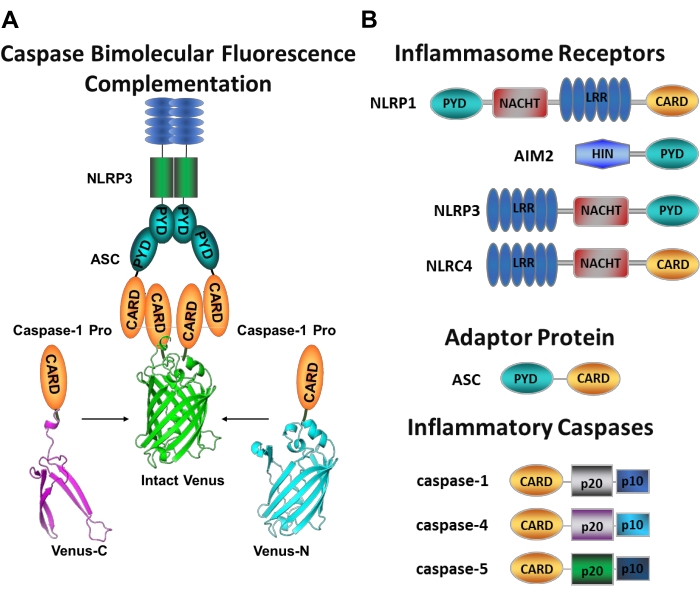
Şekil 1: İnflamatuar kaspazlar ve kaspaz-bimoleküler floresan kompleman (BiFC) testi. (A) Venüs’ün floresan olmayan her bir parçasına (Venüs-C veya Venüs-N) bağlı iki kaspaz-1 prodomaininin (C1-pro) NLRP3 aktivasyon platformuna alındığı ve Venüs’ü refere ve floresan yapmaya zorlayan kaspaz-BiFC sistemini gösteren diyagram. Bu kompleks mikroskop altında yeşil bir nokta olarak görünür ve başlatıcı kaspaz aktivasyonunda ilk adım olan inflamatuar kaspaz kaynaklı yakınlık için bir okuma görevi görür. (B) Enflamatuar bileşenlerin ve inflamatuar kaspazların etki alanı organizasyonunu gösteren şematik. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
Spesifik başlatıcı kaspaz aktivasyonunu ölçmek zordur ve görüntüleme yaklaşımlarıyla bunu yapmak için çok fazla yöntem yoktur. Kaspaz Bimoleküler Floresan Komplemanı (BiFC), doğrudan canlı hücrelerde inflamatuar kaspaz aktivasyonunu görselleştirmek için kullanılabilir (Şekil 1A)25. Bu teknik yakın zamanda insan monosit türevi makrofajlarda (MDM) kullanılmak üzere uyarlanmıştır (MDM)21. Kaspaz BiFC, dimerizasyonu kolaylaştırmak için inflamatuar kaspaz aktivasyonundaki ilk adımı, indüklenmiş yakınlığı ölçer. Fotolanabilir sarı floresan protein Venüs’ün (Venüs-C [VC]) ve Venüs-N [VN]) floresan olmayan fragmanlarına kaynaşmış CARD içeren kaspaz prodomainini kodlayan plazmidlerin ekspresyonu kullanılır. İki kaspaz prodomaini aktivasyon platformlarına alındığında veya indüklenmiş yakınlığa maruz kaldığında, Venüs’ün iki yarısı yakına getirilir ve refold ve floresan yapmaya zorlanır (bkz. Şekil 1A, B). Bu, spesifik inflamatuar kaspaz aktivasyonunun gerçek zamanlı bir okumasını sağlar.
İnsan MDM’si, tehlike sinyallerini ve patojen ürünlerini tanımlayan enflamatuar genleri ve patern tanıma reseptörlerini bolca eksprese eder. Bu, inflamatuar kaspaz yolaklarının sorgulanması için ideal bir hücre tipi sağlar. Ek olarak, periferik kandan ve hatta belirli bir hastalık durumunda inflamatuar kaspaz aktivasyonunu değerlendirmek için hasta örneklerinden türetilebilirler. Bu protokol, elektroporasyon tabanlı bir transfeksiyon yöntemi olan nükleofeksiyon kullanılarak BiFC kaspaz muhabirlerinin MDM’ye nasıl tanıtılacağını, inflamatuar kaspaz aktivasyonunu indüklemek için hücrelerin nasıl tedavi edileceğini ve mikroskopi yaklaşımlarını kullanarak aktif kaspaz komplekslerinin nasıl görselleştirileceğini açıklamaktadır. Ek olarak, bu metodoloji, bu komplekslerin moleküler bileşimini, hücre altı lokalizasyonunu, kinetik ve bu yüksek sıralı yapıların büyüklüğünü belirlemek için uyarlanabilir25,26,27.
