Caspases er en familie av cystein aspartate proteaser som kan grupperes i initiator caspases og bøddel caspases. Bøddel caspases består av caspase-3, -6 og -7. De finnes naturlig i celler som dimmere og er spaltet av initiator caspases for å utføre apoptose1. Initiator caspases inkluderer menneskelig caspase-1, -2, -4, -5, -8, -9, -10 og -12. De er funnet som inaktive zymogens (pro-caspases) som aktiveres av nærhet-indusert dimerization og stabilisert av auto-proteolytisk spalting 2,3. De inflammatoriske caspasene er en undergruppe av initiator caspases2 og omfatter caspase-1, -4, -5 og -12 hos mennesker, og caspase-1, -11 og -12 i mus 4,5. I stedet for en apoptotisk rolle, spiller de en sentral rolle i betennelse. De formidler proteolytisk prosessering og sekresjon av pro-interleukin (IL)-1β og pro-IL-18 6,7, som er de første cytokinene som slippes ut som svar på patogene inntrengere 8,9. Caspase-1 aktiveres ved rekruttering til aktiveringsplattformen; et stort molekylvektproteinkompleks kalt inflammasomet (figur 1A)10. Dimerisering av caspase-4, -5 og -11 skjer uavhengig av disse plattformene gjennom en ikke-kanonisk inflammasomvei11,12.
Kanoniske inflammasomer er cytosoliske multimeriske proteinkomplekser som består av et inflammasomsensorprotein, adapterproteinet ASC (apoptose-assosiert flekklignende protein som inneholder et CARD), og effektorproteinet caspase-110. De mest studerte kanoniske inflammasomene er den NOD-lignende reseptorfamilien som inneholder et pyrindomene (NLRP), NLRP1 og NLRP3, NLR-familien som inneholder et KORT (NLRC), NLRC4 og fraværende i melanom 2 (AIM2). Hver av dem inneholder et pyrindomene, et KORT eller begge domenene. CARD-domenet formidler samspillet mellom KORT-inneholdende caspaser og deres oppstrømsaktivatorer. Stillasmolekylet ASC, som består av et N-terminalt pyrindomene (PYD) og et C-terminalt KORTmotiv13,14, er derfor nødvendig for rekruttering av caspase-1 til NLRP110, NLRP315 og AIM216 inflammasomes.
Hvert inflammasom er oppkalt etter sitt unike sensorprotein som gjenkjenner distinkte proinflammatoriske stimuli (figur 1B). Aktivatorer av denne banen kalles kanoniske stimuli. Inflammasomer fungerer som sensorer for mikrobielle komponenter og vevsstress, og monteres for å utløse en robust inflammatorisk respons gjennom aktivering av de inflammatoriske caspases17. Inflammasome montering initierer caspase-1 aktivering for å megle modning og sekresjon av sine viktigste substrater pro-IL-1β og pro-IL-18. Denne prosessen skjer via en to-trinns mekanisme. For det første oppregulerer en priming stimulus uttrykket av visse inflammasome proteiner og pro-IL-1β gjennom aktivering av NF-κB-banen. For det andre induserer en intracellulær (kanonisk) stimulus inflammasommontering og rekruttering av procaspase-1 6,7.
Caspase-4 og caspase-5 er de menneskelige ortologene av murin caspase-1111. De aktiveres på en inflammasomuavhengig måte ved intracellulær lipopolysakkarid (LPS), et molekyl som finnes i den ytre membranen av Gram-negative bakterier 18,19,20, og ved ekstracellulært heme, et produkt av rødblodcelle hemolyse 21. Det er foreslått at LPS binder seg direkte til CARD-motivet til disse proteinene og induserer deres oligomerisering20. Aktivering av caspase-4 eller caspase-5 fremmer IL-1β-frigjøring ved å indusere en inflammatorisk form for celledød kalt pyroptose gjennom spalting av poredannende proteingassdermin D (GSDMD)18,19. I tillegg induserer efflux av kaliumioner som følge av caspase-4 og GSDMD-mediert pyroptotisk død aktivering av NLRP3 inflammasome og påfølgende aktivering av caspase-122,23. Derfor anses caspase-4, -5 og -11 som intracellulære sensorer for LPS som er i stand til å indusere pyroptose og caspase-1-aktivering som svar på spesifikke stimuli11,24.
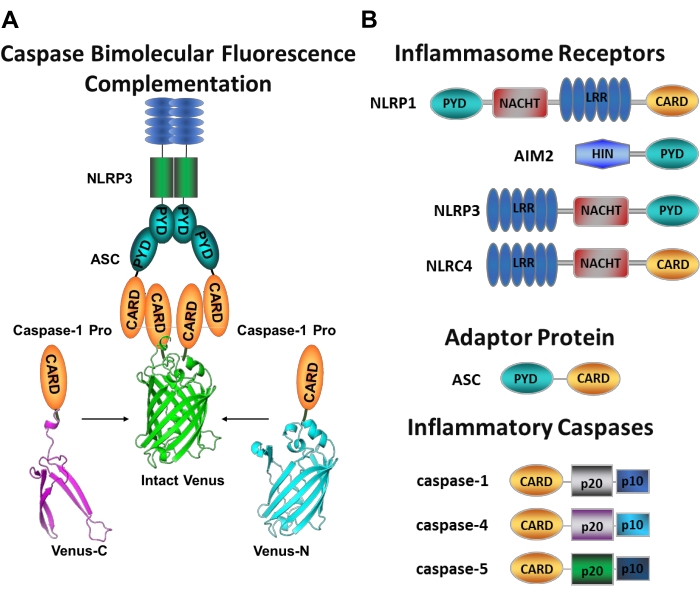
Figur 1: Inflammatoriske caspaser og caspase-bimolecular fluorescence complementation (BiFC) assay. (A) Diagram som viser caspase-BiFC-systemet, hvor to caspase-1 prodomainer (C1-pro) knyttet til hvert ikke-fluorescerende fragment av Venus (Venus-C eller Venus-N) rekrutteres til NLRP3-aktiveringsplattformen, og tvinger Venus til å refolde og fluorisere. Dette komplekset fremstår som et grønt sted under mikroskopet og fungerer som en avlesning for inflammatorisk caspase-indusert nærhet, som er det første trinnet i initiator caspase aktivering. (B) Skjematisk som viser domeneorganisasjonen av inflammasomkomponenter og inflammatoriske caspaser. Klikk her for å se en større versjon av denne figuren.
Det er vanskelig å måle spesifikk initiator caspases-aktivering, og det er ikke mange metoder tilgjengelig for å gjøre det ved avbildningsmetoder. Caspase Bimolecular Fluorescence Complementation (BiFC) kan brukes til å visualisere inflammatorisk kaspaseaktivering direkte i levende celler (figur 1A)25. Denne teknikken har nylig blitt tilpasset for bruk i humane monocytt-avledede makrofager (MDM)21. Caspase BiFC måler det første trinnet i inflammatorisk kaspaseaktivering, indusert nærhet for å lette dimerisering. Uttrykk for plasmider som koder det CARD-inneholdende caspase prodomain smeltet sammen til ikke-fluorescerende fragmenter av det fotostabile gule fluorescerende proteinet Venus (Venus-C [VC]) og Venus-N [VN]) brukes. Når de to caspase-prodomenene rekrutteres til aktiveringsplattformen eller gjennomgår indusert nærhet, blir de to halvdelene av Venus brakt i nærheten og tvunget til å brette og fluoresce (se figur 1A, B). Dette gir en sanntidsavlesning av spesifikk inflammatorisk kaspaseaktivering.
Human MDM uttrykker rikelig inflammasome gener og mønstergjenkjenningsreseptorer som identifiserer faresignaler og patogenprodukter. Dette gir en ideell celletype for avhør av inflammatoriske caspase-veier. I tillegg kan de avledes fra perifert blod og til og med fra pasientprøver for å vurdere inflammatorisk kaspaseaktivering i en bestemt sykdomstilstand. Denne protokollen beskriver hvordan man introduserer BiFC caspase-reportere i MDM ved hjelp av nukleofeksjon, en elektroporasjonsbasert transfeksjonsmetode, hvordan man behandler cellene for å indusere inflammatorisk caspase-aktivering, og hvordan du visualiserer de aktive caspase-kompleksene ved hjelp av mikroskopiske tilnærminger. I tillegg kan denne metoden tilpasses for å bestemme den molekylære sammensetningen av disse kompleksene, subcellulær lokalisering, kinetikk og størrelse på disse høyt bestilte strukturene 25,26,27.
