De fleste komplekse eukaryote proteiner gennemgår udførlige posttranslationelle modifikationer efter ekspression, hvilket kræver stærkt assisteret proteinfoldning og co-faktorer for at være funktionelle1. Produktion af store mængder opløseligt humant protein i en bakterievært er fortsat en betydelig udfordring på grund af høje omkostninger og manglen på robuste ekspressions- og oprensningsmetoder, selv for laboratorieforsøg i mindre skala2,3. MMP’er, humane endopeptidaser med stor molekylvægt, udtrykkes normalt som uopløselige inklusionslegemer, når de udtrykkes i E. coli. Ekstraktion af opløselige humane MMP’er fører ofte til en besværlig, tidskrævende opløseligheds- og genfoldningsproces4.
MMP’er har kritiske roller i både fysiologiske og patogene processer. Humane MMP’er er en familie af 23 zinkendopeptidaser, kategoriseret efter struktur og substratspecikalitet og forskelligt udtrykt på trods af et stærkt bevaret katalytisk domæne5,6. MMP’er udskilles som inaktive zymogener, reguleret via posttranslationel aktivering og deres endogene hæmmere, vævshæmmere af metalloproteinaser (TIMP’er)7,8,9,10. Selvom MMP’er oprindeligt blev anerkendt for deres rolle i ECM-omsætningen, har de også været impliceret i udvikling, morfogenese, vævsreparation og ombygning8. Dysregulering af MMP’er har især været forbundet med kræft sammen med neurodegenerative, kardiovaskulære og fibrotiske sygdomme, blandt andre sygdomme5,7.
Udviklingen af robuste MMP-produktionsmetoder i stor skala er afgørende for at sikre succesen med fremtidige undersøgelser af MMP-mekanismer gennem biokemiske og cellebaserede assays. Forskellige MMP’er er tidligere blevet udtrykt i bakterier11, herunder Hisx6-mærkede MMP’er, uden at ændre MMP-aktivitet12,13,14,15. Disse metoder inkluderer dog kedelige, lange trin, der kan være vanskelige at replikere.
Pattedyrceller kan også bruges til at udtrykke mange forskellige humane proteiner, samtidig med at de sikrer de korrekte posttranslationelle modifikationer16. Selv om pattedyrs ekspressionssystem er et ideelt valg til fremstilling af rekombinante humane proteiner med korrekte postoverensstemmende modifikationer, er de største ulemper ved denne metode indledende lave udbytter, dyre vækstmedier og reagenser, lange tidslinjer for at nå stabile ekspressionslinjer og risiko for forurening med andre arter såsom svampe eller bakterier2,11 . Desuden giver MMP-produktion i pattedyrs cellelinjer urenheder fra associerede cellulære proteiner såsom TIMP’er eller fibronectiner11. I modsætning til den langsomme cellevækst, der observeres i pattedyrceller, tilbyder bakterieekspressionssystemet storskala proteinproduktion på kort tid sammen med enklere medie- og vækstkrav. På grund af manglen på andre associerede cellulære proteiner (dvs. TIMP’er) i bakterielle ekspressionssystemer udsættes aktive MMP’er i højere koncentrationer imidlertid for nedbrydning gennem autoproteolyse, hvilket resulterer i dårligt MMP-udbytte17.
Dette papir beskriver en detaljeret metode til bakteriel ekspression, oprensning og aktivering af rekombinant Hisx6-pro-MMP-3cd ved hjælp af E. coli som ekspressionsvært på grund af dets overkommelige priser, enkelhed og succes med at producere højere udbytter af MMP’er2,3,18. Da E. coli mangler proteinfoldemaskineri og posttranslationel behandling, der kræves til rekombinante MMP’er og andre komplekse proteiner, er mange E. coli-stammer blevet konstrueret til at overvinde disse begrænsninger, hvilket gør E. coli til en mere egnet vært til ekspression af rekombinant human MMP-3cd,19,20 . For eksempel forbedrer R2DP-stammen, der anvendes i denne undersøgelse, eukaryot ekspression ved at levere et chloramphenicolresistent plasmid indeholdende kodoner, der sjældent anvendes i E. coli.
Som beskrevet i denne protokol ekstraheres hisx6-pro-MMP-3 katalytiske domæneproteiner (MMP-3cd) efter overekspression af relativt rene inklusionslegemer fra pET-3a-vektoren (figur 1) i R2DP-celler4. Hisx6-pro-MMP-3cd3,19 blev renset ved hjælp af affinitetsmærkekromatografi. Ved omfoldning og dialyse blev pro-MMP-3cd (zymogen) aktiveret af 4-aminophenylmercuric acetat (APMA), og SDS-PAGE-analyse anvendes til at evaluere udbytter og behovet for yderligere oprensning5,21. Denne protokol beskriver ekspression, oprensning og aktivering af opløselig mMP-3cd som et eksempel. Det kan dog også anvendes som vejledning til ekspression af andre MMP’er og humane proteaser med lignende ekspression og aktiveringsmekanismer (figur 2). For andre proteiner end MMP-3cd rådes læseren til at bestemme optimale buffersammensætninger og metoder til deres målprotein, inden denne protokol forsøges.
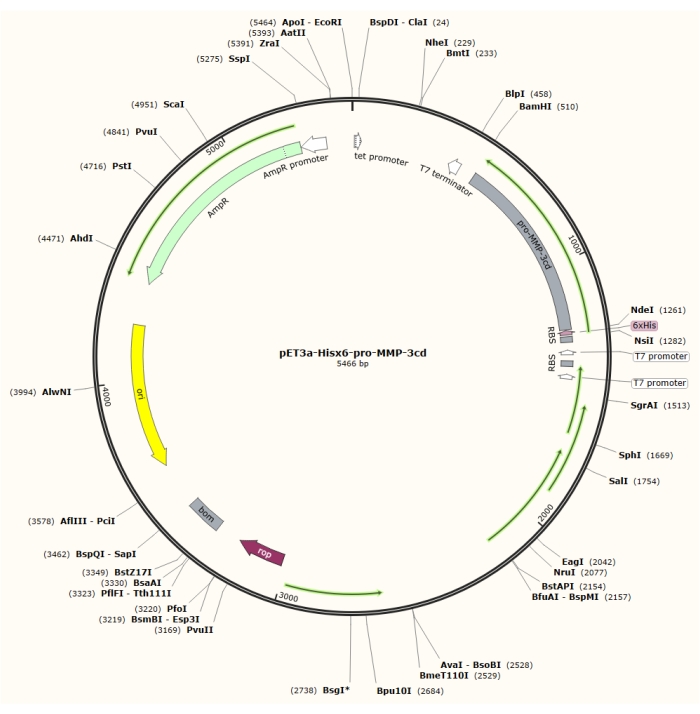
Figur 1: Plasmidkort over pET-3a-Hisx6-pro-MMP-3cd-plasmidet. PET-3a-vektoren indbefatter et ampicillinresistensgen. En N-terminal Hisx6-tag-sekvens klones ind i den pET-3a-baserede vektor, herunder pro-MMP-3cd, for at give pET-3a-Hisx6-pro-MMP-3cd-konstruktionen under kontrol af T7-promotor mellem BamHI- og NdeI-begrænsningssteder. Klik her for at se en større version af denne figur.
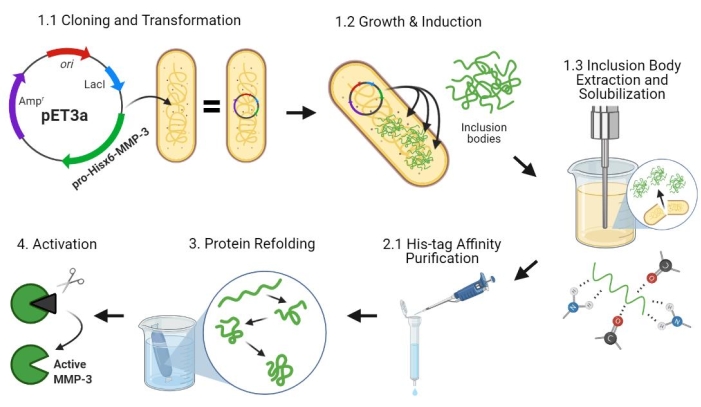
Figur 2: Bakteriel ekspression af pro-MMP-3cd, oprensning, omfoldning og aktivering. 1.1: pET-3a-Hisx6-pro-MMP-3cd plasmid blev omdannet til BL21 (DE3) eller R2DP-celler. 1.2: Pro-MMP-3cd-proteinekspression blev induceret under anvendelse af IPTG. 1.3: Kemisk lysis og sonikering bruges til at ekstrahere Hisx6-pro-MMP-3cd-proteiner, der hovedsageligt er uopløselige og findes i inklusionsorganerne. Urea blev brugt til at denaturere og opløse protein fra inklusionslegemer. 2.1. Denatureret Hisx6-pro-MMP-3cd-protein blev oprenset via affinitetskromatografirensning. 3. Den eluerede Hisx6-pro-MMP-3cd blev langsomt foldet om under dialyse gennem gradvis fjernelse af urinstof fra bufferen. 4. Endelig blev genfoldet MMP-3cd-protein aktiveret ved hjælp af APMA ved at fjerne det N-terminale propeptiddomæne. APMA fjernes senere fra opløsningen ved afsaltning. Tallene svarer til protokolafsnit, der beskriver disse trin. Forkortelser: MMP-3cd = Matrix metalloproteinase-3 katalytisk domæne; APMA = 4-aminophenylmercuric acetat. Klik her for at se en større version af denne figur.
