Following the above-mentioned methods, 5 different strains of P. lacuna were isolated from rockpools and sequenced (Figure 1 and Table 1). All cultures were sterile after ~1 year of subculturing except P. lacuna HE10JO. This strain is still contaminated with Marivirga atlantica, a marine bacterium. During subsequent Helgoland excursions, other filamentous cyanobacteria were isolated from rock pools, which are different from P. lacuna and need to be characterized.
Several DNA extraction and purification methods were tested for P. lacuna. The best results were obtained with an optimized CTAB method as described above. DNA yields were 310 ± 50 µg/mL, OD 260 nm/OD 280 nm was 1.7 ± 0.03, and OD 260 nm/OD 230 nm was 0.78 ± 0.04 (n = 17). Genome sequencing showed that the DNA of all strains was slightly different, as expected (Table 1). Core protein sequences showed a maximum difference of 0.04% (Table 2). Although all draft genomes were incomplete, one can assume that >98% of the genome of HE10JO30 was sequenced. This estimation is based on the number of incomplete open reading frames. Partial protein sequences could be easily identified after RAST annotation of HE10DO and HE10JO. In HE10JO, 60 proteins out of ~4,500 had a missing N- or C-terminal sequence. The genome sequences can be found in the supplement (Supplemental File 3, Supplemental File 4, Supplemental File 5, Supplemental File 6, and Supplemental File 7).
Interestingly, strains of the same species were isolated from two islands, Helgoland and Giglio. The linear distance between both islands is 1,400 km. There must be a link between both places, e.g., by ships via the sea or, more likely, by migratory birds. Many bird species can be found on both islands, and many of them are migratory birds. The diversity within P. lacuna strains of one island was greater than between the closest Helgoland and Giglio strains (Table 2). This indicates an intense exchange between both places.
The natural transformation was tested with HE10DO as the major strain and with HE10JO. The present protocol is more straightforward than the protocol described earlier12 because of the reduced number of washing steps and fewer transfer steps after transformation. This new method is continuously used in the laboratory; ~15 successful transformations were achieved.
The KanR resistance cassette was usually integrated into the homologous site defined by the adjacent regions, as shown by PCR using inner and outer primers. Like most cyanobacteria, P. lacuna is polyploid. It can have more than 100 chromosomes per cell12. A PCR test with outer primers ~1 week after the transformation typically has 2 bands on the electrophoresis gel, one with the size of the wild-type band and one slower migrating band that indicates the insertion of the resistance cassette (Figure 4). The double band indicates that only a subfraction of the chromosomes contains the insertion. After 4 weeks of selection on kanamycin, segregation is usually complete, and only one large PCR band appears on gels. However, in the case of the transformation with pMH1 (see below), segregation was complete after more than 3 months.
The vectors pAK1, pAK2, pAK3, and pMH1 were constructed for tests on sfGFP expression. In pAK1, pAK2, and pAK3, the sfGFP gene is under the control of the cpc560, A2813, and psbA2 promoters, respectively. These promoters are from Synechocystis sp. PCC 6803 or Synechococcus sp. PCC 700231. For the construction of these vectors, the sfGFP promoter and terminator sequences were taken from vectors used for the transformation of Synechococcus sp. PCC 700231. The relevant sequences were integrated into the homologous chwA (sc_7_37) site of pFN1 (or pFN_7_37_KanR15). The pMH1 expression vector was constructed by DNA synthesis using P. lacuna sequences as templates (Supplemental File 6). The cpcB–cpcA (phycocyanin ß and phycocyanin α) sequences of P. lacuna are serially arranged. A 100 bp intergenic region separates both coding regions. The synthetic sequence contained this endogenous cpcB–cpcA sequence and the cpcB promoter. The sfGFP and KanR cassette is placed just 3' of the cpcB stop codon (5' of cpcA). The entire synthetic sequence with cpcB promoter, cpcB, sfGFP, KanR, cpcA (5' to 3') is cloned into pUC19. A map is shown in Figure 5. More details on the cloning of pAK1, pAK2, and pAK3 and the complete sequence of pMH1 are given in Supplemental File 1.
All 4 transformants (with pAK1, pAK2, pAK3, and pMH1) expressed GFP; all fluorescence levels were above the background fluorescence of wild-type filaments (Figure 6). The pMH1 transformants with incomplete segregation revealed a GFP signal that was very variable between the filaments. The fluorescence signal was evenly distributed when segregation was complete (Figure 6E). The microscope signals of pAK1, pAK2, and pAK3 transformants were similar but ~5x weaker than that of pMH1 (Figure 6E).
The established cryoconservation method is based on a method that was established for E. coli. When 2 washing steps were performed for glycerol removal after thawing, 15 out of 15 P. lacuna samples survived (Table 3). This protocol could also be used for Synechocystis PCC 6803, but only with 2 washing steps and not with 1 (Table 3).
Another feature of Oscillatoriales filaments is their motility: P. lacuna filaments move continuously on surfaces (Figure 7) and in a liquid medium (Figure 8). Both kinds of motion can be studied easily in Petri dishes without or with agar medium. Time-lapse recording is required because movement on agar is slow. Filaments move towards the light cone if a light beam comes from below (Figure 3). The effects of light intensity, wavelength, and time can be easily studied with a simple setup. The photoreceptors of this effect are not yet clear. Possible candidates can be addressed with knockout mutants. The mechanism underlying how the filaments find the light is also unclear. For this question, an infrared system is required to record the filaments during their movement from darkness to light.
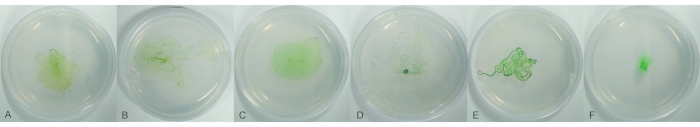
Figure 1: Strains of Phormidium lacuna collected from Helgoland and Giglio. Filaments are propagated for 11 days on f/2 agar in 6 cm Petri dishes. (A) strain GI08AO; (B) strain GI08IO; (C) strain GI09CO; (D) strain HE10DO; (E) strain HE10JO; (F) strain HE15M2G1. Please click here to view a larger version of this figure.
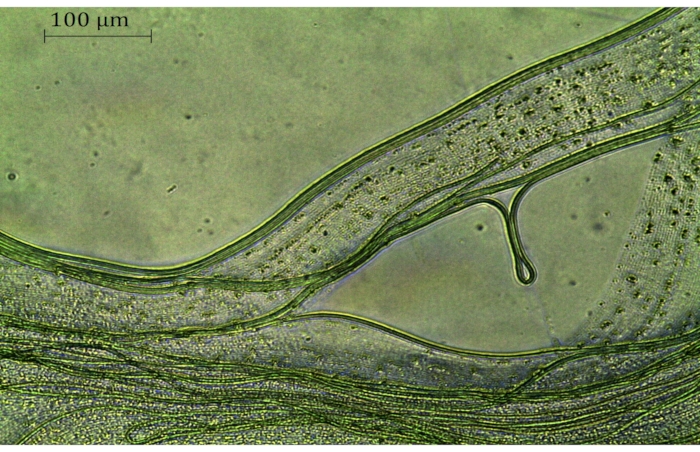
Figure 2: Phormidium lacuna filaments 5 weeks after transformation. The sfGFP expression vector pMH1 was used; selection occurred on f/2+ medium with 120 µg/mL kanamycin. The greenish filaments are resistant and alive; other filaments have died. Scale bar = 100 µm. Please click here to view a larger version of this figure.
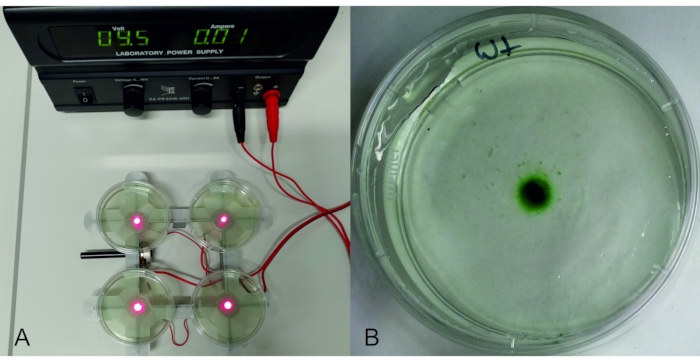
Figure 3: Phototaxis experiment. Left: LED holder with 4 red LEDs, connected to an adjustable power supply. On top of each LED, there is a 6 cm Petri dish with 8 mL of a Phormidium lacuna culture. Right: Petri dish with P. lacuna after 2 days on the red LED (15 µmol m-2 s-1). Abbreviation: LED = light-emitting diode. Please click here to view a larger version of this figure.
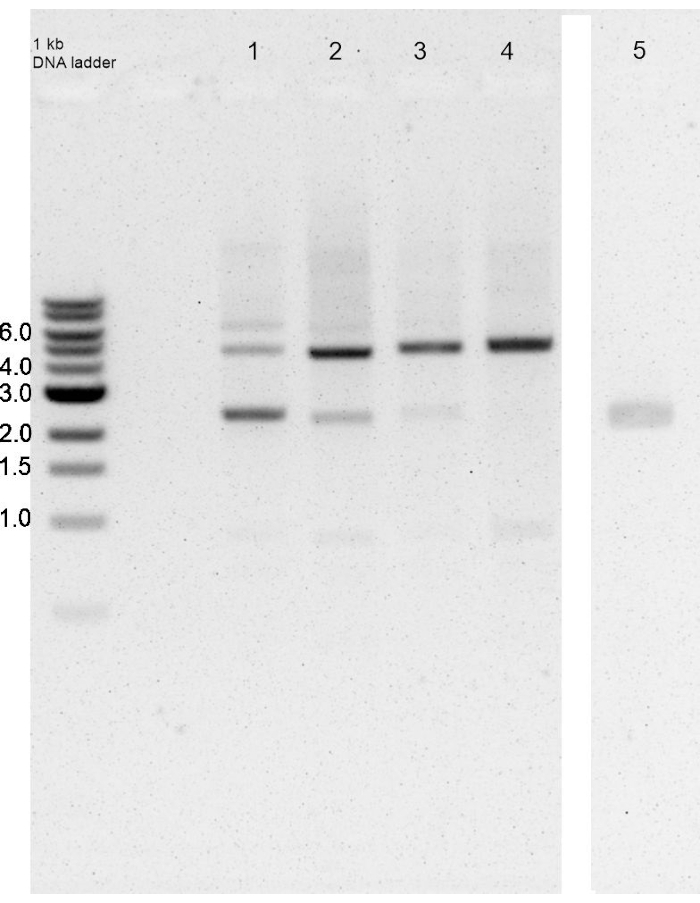
Figure 4: Integration and segregation of insert after transformation of Phormidium lacuna with pAK1. PCR with outer primers. The expected sizes of the product without and with insert are 2371 and 5016 bp, respectively. Left lane: marker, lanes 1, 2, 3, 4: PCR products of filaments 7 days, 11 days, 14 days, and 17 days after the isolation of a resistant filament (4 weeks after transformation), respectively. Lane 5: PCR product of wild-type (from a different gel). In the 7 day sample, the insert is present in a small fraction of the chromosomes. This fraction increases until 17 days, where no wild-type band is visible, i.e., segregation is complete. Please click here to view a larger version of this figure.
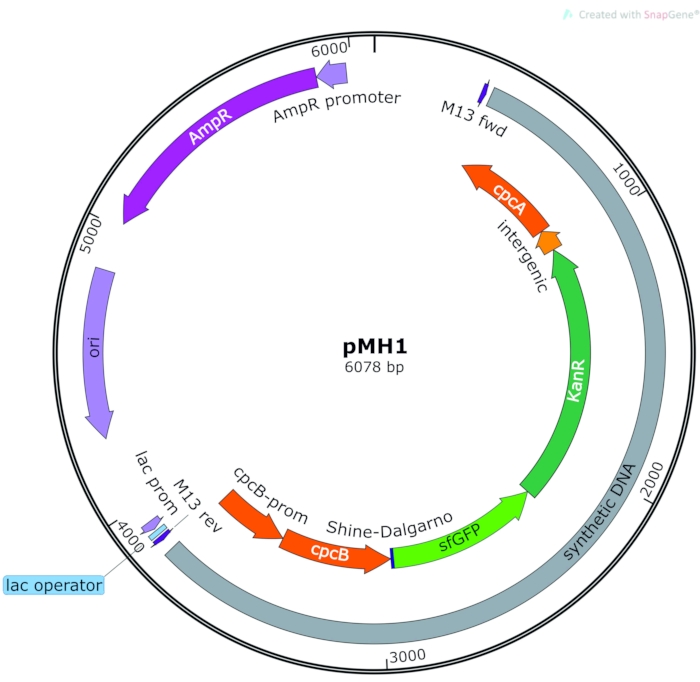
Figure 5: Vector for sfGFP expression under the control of endogenous cpcB promoter. Orange: Phormidium lacuna homologous sequence, violet/blue: pUC-19 vector backbone, green: insert with sfGFP and KanR. Abbreviations: sfGFP = superfolder green fluorescent protein; KanR = kanamycin resistance. Please click here to view a larger version of this figure.
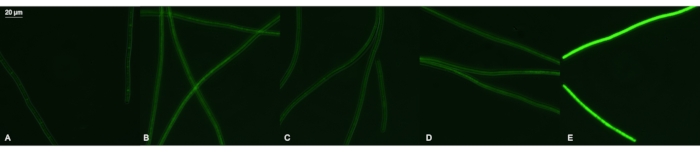
Figure 6: Expression of sfGFP in Phormidum lacuna. Fluorescence images of P. lacuna wild-type filaments (A) and after transformation with pAK1 (B), pAK2 (C), pAK3 (D), and pMH1 (E). In pMH1, the sfGFP gene is placed 3' of the phycocyanin ß gene and therefore driven by the endogenous cpcß promoter; in the other cases, sfGFP is driven by cpc560, A2813, or psbA2s promoters from Synechocystis PCC 6803, respectively. The fluorescence settings are specific for GFP; all images were recorded with the same integration time and optical settings. Please click here to view a larger version of this figure.
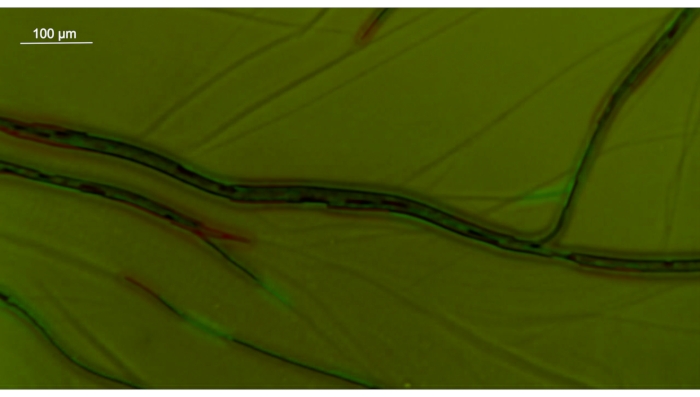
Figure 7: Merged image of Phormidium lacuna on agar surface at 4x magnification. The first image is presented in red, the second (taken 1 min later) in green. Note also the traces on the agar. Scale bar = 100 µm. Please click here to view a larger version of this figure.

Figure 8: Merged image of Phormidium lacuna in liquid medium. The time interval between both images was 10 s. The first image is printed in red; the second is printed in green. Comparing both colors shows the movement within 10 s. Scale bar = 100 µm. Please click here to view a larger version of this figure.
| strain | HE10JO | HE10DO | GI08AO | GI09CO | HE15M2G1 |
| Contigs | 104 | 174 | 218 | 102 | 154 |
| total bp | 48,19,017 | 47,88,491 | 47,78,775 | 36,69,922 | 45,98,395 |
Table 1: Phormidium lacuna strains.
| Gi09CO | HE10DO | HE10JO | HE15M2G1 | |
| Gi08AO | 42 | 40 | 0 | 42 |
| Gi09CO | 2 | 42 | 0 | |
| HE10DO | 40 | 2 | ||
| HE10JO | 42 |
Table 2. Amino acid differences between strains in sequences of 20 core proteins with 10,876 amino acids.
| Cell density OD 750 nm | 1 | 1 | 3 | 3 | 5 | 5 | 7 | 7 |
| Washes | 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 |
| Synechocystis PCC 6803 | 2/5 | 5/5 | 1/4 | 4/4 | 0/4 | 4/4 | 0/4 | 3/4 |
| Phormidium lacuna HE10DO | 4/4 | 4/4 | 4/4 | 4/4 | 3/ 4 | 4/4 | 3/3 | 3/3 |
Table 3: Cryoconservation trials with Synechocystis PCC 6803 and Phormidium lacuna HE10DO cyanobacteria. The first number shows the number of cultures that survived after freezing/thawing; the second number shows the total trials.
Supplemental Video S1: Movement of Phormidium lacuna filaments in liquid solution, without time-lapse. Please click here to download this Video.
Supplemental Video S2: Movement of Phormidium lacuna filaments on agar surface, with time-lapse. Please click here to download this Video.
Supplemental File 1: Cloning of vectors for transformation of Phormidium lacuna. List of transformation vectors; list of primers for cloning; sequence of pMH1 in gb format. Please click here to download this File.
Supplemental File 2: Shell scripts (sh) for Raspberry Pi minicomputer. Please click here to download this File.
Supplemental File 3: DNA sequence of HE152G1. Please click here to download this File.
Supplemental File 4: DNA sequence of GI08AO. Please click here to download this File.
Supplemental File 5: DNA sequence of GI09CO. Please click here to download this File.
Supplemental File 6: DNA sequence of HE10DO. Please click here to download this File.
Supplemental File 7: DNA sequence of HE10JO. Please click here to download this File.
