इस विधि का लक्ष्य यह जांचना है कि रेटिना इनपुट ऑप्टिक टेक्टम के विकास और विकास को कैसे प्रभावित करता है, जो ज़ेब्राफ़िश मस्तिष्क में दृश्य प्रसंस्करण केंद्र है। एक आंख को हटाकर और फिर ऑप्टिक टेक्टम के दो पक्षों की तुलना करके, एक ही नमूने के भीतर टेक्टल परिवर्तनों को देखा जा सकता है और सामान्यीकृत किया जा सकता है, जिससे कई नमूनों में तुलना की जा सकती है। इस तकनीक के साथ संयुक्त आधुनिक आणविक दृष्टिकोण दृश्य प्रणाली के विकास और विकास के अंतर्निहित तंत्र में अंतर्दृष्टि प्राप्त करेंगे, साथ ही साथ अक्षीय अध: पतन और पुनर्जनन भी।
संवेदी प्रणालियां – दृश्य, श्रवण, और सोमाटोसेंसरी – बाहरी अंगों से जानकारी इकट्ठा करती हैं और उस जानकारी को केंद्रीय तंत्रिका तंत्र को रिले करती हैं, जिससे मिडब्रेन 1,2 में बाहरी दुनिया के “मानचित्र” उत्पन्न होते हैं। दृष्टि लगभग सभी कशेरुकियों के लिए प्रमुख संवेदी रूपरेखा है, जिसमें कई मछलियां भी शामिल हैं। रेटिना, आंखों में तंत्रिका ऊतक, एक न्यूरोनल सर्किट के साथ जानकारी एकत्र करता है जिसमें मुख्य रूप से फोटोरिसेप्टर, द्विध्रुवी कोशिकाएं और रेटिना गैंग्लियन कोशिकाएं (आरजीसी), रेटिना के प्रक्षेपण न्यूरॉन्स शामिल होते हैं। आरजीसी में लंबे अक्षतंतु होते हैं जो रेटिना की आंतरिक सतह पर ऑप्टिक तंत्रिका सिर तक अपना रास्ता ढूंढते हैं, जहां वे मस्तिष्क के माध्यम से एक साथ यात्रा करते हैं, अंततः पृष्ठीय मिडब्रेन में दृश्य प्रसंस्करण केंद्र में समाप्त हो जाते हैं। इस संरचना को मछली और अन्य गैर-स्तनधारी कशेरुकियों में ऑप्टिक टेक्टम कहा जाता है और स्तनधारियों में बेहतर कोलिकुलस के लिए समरूप है।
ऑप्टिक टेक्टम पृष्ठीय मध्यस्तिष्क में एक द्विपक्षीय सममित बहुस्तरीय संरचना है। ज़ेब्राफ़िश और अधिकांश अन्य मछलियों में, ऑप्टिक टेकटम का प्रत्येक लोब पूरी तरह से contralateral आंख से दृश्य इनपुट प्राप्त करता है, जैसे कि बाएं ऑप्टिक तंत्रिका दाएं टेक्टल लोब में समाप्त हो जाती है और दाईं ऑप्टिक तंत्रिका बाएं टेक्टल लोब4 (चित्रा 1) में समाप्त हो जाती है। अपने स्तनधारी समकक्ष की तरह, बेहतर कोलिकुलस, ऑप्टिक टेकटम अन्य संवेदी इनपुट के साथ दृश्य जानकारी को एकीकृत करता है, जिसमें ऑडिशन और सोमाटोसेंसेशन शामिल हैं, दृश्य ध्यान और आंखों के आंदोलनों में बदलाव को नियंत्रित करते हैं जैसे कि सैकेड्स 1,5,6। हालांकि, स्तनधारी बेहतर कोलिकुलस के विपरीत, ऑप्टिक टेकटम लगातार टेक्टल लोब के औसत दर्जे के और पुच्छल किनारों के पास एक विशेष स्टेम सेल आला से नए न्यूरॉन्स और ग्लिया उत्पन्न करता है जिसे टेक्टल प्रसार क्षेत्र7 कहा जाता है। ऑप्टिक टेकटम और केंद्रीय तंत्रिका तंत्र के अन्य क्षेत्रों में प्रोलिफेरेटिव पूर्वजों का रखरखाव, भाग में, ज़ेबराफ़िश 8 में प्रलेखित उल्लेखनीय पुनर्योजी क्षमता में योगदान देताहै।
अंधे या एक आंखों वाली मछलियों के दिमाग की जांच करने वाले पिछले काम से पता चला कि ऑप्टिक टेक्टम का आकार रेटिना इनर्वेशन की मात्रा के लिए सीधे आनुपातिक है जोइसे 9,10,11 प्राप्त करता है। वयस्क गुफा मछली में, जिनकी आंखें प्रारंभिक भ्रूणजनन में पतित होती हैं, ऑप्टिक टेकटम निकटता से संबंधित, दृष्टिहीन सतह मछली9 की तुलना में काफी छोटा होता है। गुफा मछली आंख अध: पतन भ्रूणजनन के दौरान एक सतह मछली से एक लेंस के साथ अंतर्जात लेंस को बदलकर अवरुद्ध किया जा सकता है। जब इन एक आंखों वाली गुफा मछलियों को वयस्कता में पाला जाता है, तो इनरवेटेड टेक्टल लोब में गैर-इनरवेटेड टेक्टल लोब 9 की तुलना में लगभग 10% अधिक कोशिकाएं होतीहैं। इसी तरह, लार्वा किलीफ़िश में जो एक ही व्यक्ति के भीतर विभिन्न आकारों की आंखों को उत्पन्न करने के लिए रासायनिक उपचार के साथ इनक्यूबेट किए गए थे, अधिक संरक्षण के साथ टेक्टम का पक्ष बड़ा था और इसमें अधिक न्यूरॉन्स10 शामिल थे। वयस्क सुनहरी मछली में ऑप्टिक तंत्रिका क्रश प्रयोगों से सबूत इंगित करता है कि संरक्षण प्रसार को बढ़ावा देता है, टेक्टल सेल प्रसार के साथ कम हो जाता है जब इनरवेशन बाधित हो गया था11।
इन शास्त्रीय अध्ययनों की पुष्टि और विस्तार करते हुए, कई हालिया रिपोर्टें डेटा प्रदान करती हैं जो सुझाव देती हैं कि इनरवेशन के जवाब में प्रसार को कम से कम भाग में, BDNF-TrkB Pathway12,13 द्वारा संशोधित किया जाता है। ऑप्टिक टेक्टम विकास और विकास के बारे में कई खुले प्रश्न बने हुए हैं, जिसमें यह भी शामिल है कि एक विकासशील संवेदी प्रणाली चोट और अक्षतंतु अध: पतन के साथ कैसे सामना करती है, जो सेलुलर और आणविक संकेत ऑप्टिक टेकटम विकास को विनियमित करने के लिए रेटिना इनपुट को सक्षम करते हैं, जब ये तंत्र सक्रिय हो जाते हैं, और क्या इनरवेशन-लिंक्ड प्रसार और भेदभाव रेटिना और इसके लक्ष्य ऊतक को विकास दरों का समन्वय करने और सटीक रेटिनोटोपिक मैपिंग सुनिश्चित करने में सक्षम बनाते हैं। इसके अलावा, गतिविधि-निर्भर विकास के बारे में बहुत बड़े प्रश्न हैं जिन्हें सर्जिकल दृष्टिकोण के साथ ज़ेब्राफ़िश दृश्य प्रणाली से पूछताछ करके संबोधित किया जा सकता है जैसे कि नीचे वर्णित एक।
सेलुलर और आणविक तंत्र की जांच करने के लिए जिसके द्वारा तंत्रिका गतिविधि, विशेष रूप से दृश्य इनपुट से, सेल अस्तित्व और प्रसार को बदल देती है, वर्णित दृष्टिकोण सीधे व्यक्तिगत ज़ेब्राफ़िश लार्वा के भीतर आंतरिक और व्युत्पन्न टेक्टल लोब (चित्रा 1) की तुलना करता है। यह विधि ऑप्टिक टेकटम में आरजीसी अक्षतंतु अध: पतन के प्रलेखन और पुष्टि के लिए अनुमति देती है कि माइटोटिक कोशिकाओं की संख्या इनरवेशन के साथ सहसंबंधित है।
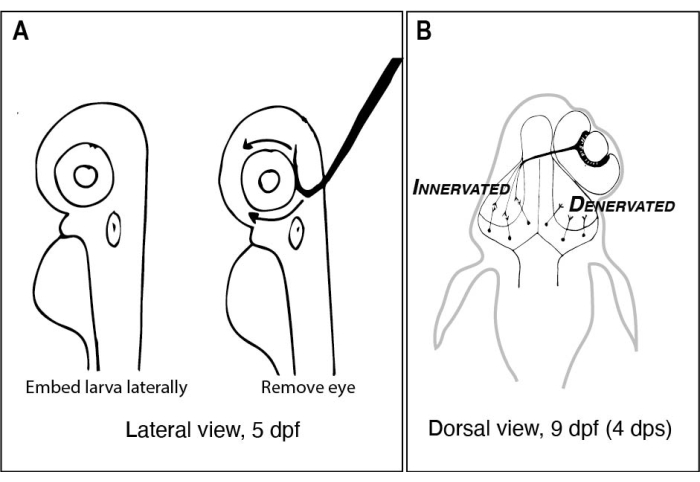
चित्र 1: एकतरफा आंख हटाने से पहले और बाद में ज़ेब्राफ़िश लार्वा के स्केच। (A) विच्छेदन माइक्रोस्कोप के नीचे देखे गए 5 dpf लार्वा का आरेखण। प्रत्येक लार्वा कम पिघलने-बिंदु agarose में एम्बेडेड है और एक तेज, झुका हुआ टिप के साथ एक टंगस्टन सुई से पहले पार्श्व रूप से उन्मुख है, ऊपर का सामना कर रही आंख (इस उदाहरण में बाईं आंख) का सामना कर रही आंख को स्कूप करने के लिए उपयोग किया जाता है। (बी) ए में दर्शाई गई सर्जरी के परिणामस्वरूप 9 डीपीएफ लार्वा के पृष्ठीय दृश्य का ड्राइंग। दाईं आंख से केवल तीन अत्यधिक योजनाबद्ध आरजीसी अक्षतंतुओं को बाएं टेक्टल लोब में न्यूरॉन्स के साथ विघटित और कनेक्ट करते हुए दिखाया गया है। संक्षेप: dpf = निषेचन के बाद के दिन; डीपीएस = सर्जरी के बाद के दिन; RGC = रेटिना गैंग्लियन कोशिकाएं। कृपया इस आंकड़े का एक बड़ा संस्करण देखने के लिए यहाँ क्लिक करें.
