الهدف من هذه الطريقة هو التحقيق في كيفية تأثير مدخلات الشبكية على نمو وتطور التكتوم البصري ، وهو مركز المعالجة البصرية في دماغ الزرد. من خلال إزالة عين واحدة ثم مقارنة جانبي التكتوم البصري ، يمكن ملاحظة التغيرات التكتالية داخل نفس العينة وتطبيعها ، مما يتيح المقارنة عبر عينات متعددة. سوف تسفر الأساليب الجزيئية الحديثة جنبا إلى جنب مع هذه التقنية عن رؤى ثاقبة حول الآليات الكامنة وراء نمو النظام البصري وتطوره ، بالإضافة إلى الانحطاط والتجديد المحوري.
تجمع الأنظمة الحسية – البصرية والسمعية والحسية الجسدية – المعلومات من الأعضاء الخارجية وتنقل تلك المعلومات إلى الجهاز العصبي المركزي ، وتولد “خرائط” للعالم الخارجي عبر الدماغ المتوسط 1,2. الرؤية هي الطريقة الحسية السائدة لجميع الفقاريات تقريبا ، بما في ذلك العديد من الأسماك. تجمع شبكية العين، وهي النسيج العصبي في العين، المعلومات مع دائرة عصبية تتكون في المقام الأول من المستقبلات الضوئية، والخلايا ثنائية القطب، وخلايا العقدة الشبكية (RGCs)، وهي الخلايا العصبية الإسقاط في شبكية العين. تحتوي RGCs على محاور طويلة تجد طريقها عبر السطح الداخلي للشبكية إلى رأس العصب البصري ، حيث تبهر وتنتقل معا عبر الدماغ ، وتنتهي في النهاية في مركز المعالجة البصرية في الدماغ المتوسط الظهري. ويسمى هذا الهيكل التكتوم البصري في الأسماك وغيرها من الفقاريات غير الثدييات وهو متجانس مع كوليكولوس متفوقة في الثدييات3.
التكتوم البصري هو بنية متعددة الطبقات متماثلة ثنائيا في الدماغ المتوسط الظهري. في أسماك الزرد ومعظم الأسماك الأخرى ، يتلقى كل فص من التكتوم البصري مدخلات بصرية فقط من العين المقابلة ، بحيث ينتهي العصب البصري الأيسر في الفص التكتال الأيمن وينتهي العصب البصري الأيمن في الفص التكتالي الأيسر4 (الشكل 1). مثل نظيره الثدييات ، كوليكولوس المتفوق ، يدمج التكتوم البصري المعلومات البصرية مع المدخلات الحسية الأخرى ، بما في ذلك الاختبار والإحساس الجسدي ، والتحكم في التحولات في الانتباه البصري وحركات العين مثل saccades 1,5,6. ومع ذلك ، على عكس كوليكولوس الثدييات العليا ، فإن التكتوم البصري يولد باستمرار خلايا عصبية ودبقية جديدة من مكانة الخلايا الجذعية المتخصصة بالقرب من الحواف الإنسية والذيلية للفصوص التكتالية التي تسمى منطقة الانتشار التكتالي7. تساهم صيانة السلف التكاثرية في التكتوم البصري ومناطق أخرى من الجهاز العصبي المركزي ، جزئيا ، في القدرة التجديدية الرائعة الموثقة في الزرد8.
كشفت الأبحاث السابقة التي فحصت أدمغة الأسماك العمياء أو ذات العين الواحدة أن حجم التكتوم البصري يتناسب طرديا مع كمية تعصيب الشبكية التي يتلقاها9،10،11. في أسماك الكهوف البالغة ، التي تتدهور عيونها في التكوين الجنيني المبكر ، يكون التكتوم البصري أصغر بشكل ملحوظ من الأسماك السطحية المبصرة ذات الصلة الوثيقة9. يمكن منع تنكس عين سمكة الكهف عن طريق استبدال العدسة الداخلية بعدسة من سمكة سطحية أثناء التكوين الجنيني. عندما يتم تربية أسماك الكهوف ذات العين الواحدة هذه حتى مرحلة البلوغ ، يحتوي الفص التكتال المعصب على خلايا أكثر بنسبة 10٪ تقريبا من الفص التكتال غير المعصب9. وبالمثل ، في أسماك اليرقات اليرقات التي تم احتضانها بعلاجات كيميائية لتوليد عيون بأحجام مختلفة داخل نفس الفرد ، كان جانب التكتوم مع المزيد من التعصيب أكبر ويحتوي على المزيد من الخلايا العصبية10. تشير الأدلة المستقاة من تجارب سحق العصب البصري في الأسماك الذهبية البالغة إلى أن التعصيب يعزز الانتشار ، مع انخفاض تكاثر الخلايا التكتالية عندما تعطل التعصيب11.
وتأكيدا لهذه الدراسات الكلاسيكية وتوسيعها، تقدم العديد من التقارير الحديثة بيانات تشير إلى أن الانتشار استجابة للتعصيب يتم تعديله، جزئيا على الأقل، من خلال مسار BDNF-TrkB12,13. لا تزال هناك العديد من الأسئلة المفتوحة حول نمو التكتوم البصري وتطوره ، بما في ذلك كيفية تعامل النظام الحسي النامي مع الإصابة وانحطاط المحور العصبي ، والإشارات الخلوية والجزيئية التي تمكن مدخلات الشبكية من تنظيم نمو التكتوم البصري ، ومتى تصبح هذه الآليات نشطة ، وما إذا كان الانتشار والتمايز المرتبطان بالتعصيب يمكنان شبكية العين وأنسجتها المستهدفة من تنسيق معدلات النمو وضمان رسم خرائط دقيقة للشبكية الشبكية. بالإضافة إلى ذلك ، هناك أسئلة أكبر بكثير حول التطور المعتمد على النشاط والتي يمكن معالجتها عن طريق استجواب النظام البصري لأسماك الزرد باستخدام الأساليب الجراحية مثل تلك الموضحة أدناه.
للتحقيق في الآليات الخلوية والجزيئية التي يغير بها النشاط العصبي ، وتحديدا من المدخلات البصرية ، بقاء الخلايا وانتشارها ، يقارن النهج الموصوف مباشرة الفصوص التكتالية المعصبة والمشوهة (الشكل 1) داخل يرقات الزرد الفردية. تسمح هذه الطريقة بتوثيق تنكس محور RGC في التكتوم البصري وتأكيد أن عدد الخلايا الانقسامية يرتبط بالتعصيب.
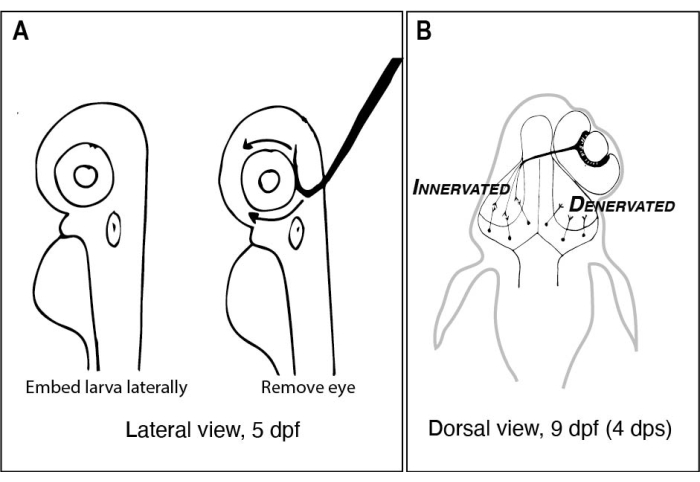
الشكل 1: رسومات ليرقات الزرد قبل وبعد إزالة العين من جانب واحد . (أ) رسم يرقات 5 dpf كما ينظر إليها تحت المجهر تشريح. يتم تضمين كل يرقة في الأغاروز منخفض الانصهار وتوجيهها أفقيا قبل استخدام إبرة التنغستن ذات الطرف الحاد المدمن على المخدرات لإخراج العين متجهة لأعلى (العين اليسرى في هذا المثال). (ب) رسم المنظر الظهري ليرقة 9 dpf الناتجة عن الجراحة الموضحة في A. تظهر ثلاثة محاور RGC عالية التخطيط فقط من العين اليمنى وهي تزيل اللفافة وتتصل بالخلايا العصبية في الفص التكتالي الأيسر. الاختصارات: dpf = أيام بعد الإخصاب ؛ dps = أيام ما بعد الجراحة; RGC = خلايا العقدة الشبكية. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
