Het doel van deze methode is om te onderzoeken hoe retinale input de groei en ontwikkeling van het optische tectum, het visuele verwerkingscentrum in het zebravisbrein, beïnvloedt. Door één oog te verwijderen en vervolgens de twee zijden van het optische tectum te vergelijken, kunnen tectale veranderingen binnen hetzelfde monster worden waargenomen en genormaliseerd, waardoor vergelijking tussen meerdere monsters mogelijk is. Moderne moleculaire benaderingen in combinatie met deze techniek zullen inzicht geven in de mechanismen die ten grondslag liggen aan de groei en ontwikkeling van visuele systemen, evenals axonale degeneratie en regeneratie.
Sensorische systemen – visueel, auditief en somatosensorisch – verzamelen informatie van externe organen en geven die informatie door aan het centrale zenuwstelsel, waardoor “kaarten” van de externe wereld in de middenhersenen worden gegenereerd 1,2. Visie is de dominante sensorische modaliteit voor bijna alle gewervelde dieren, inclusief veel vissen. Het netvlies, het neurale weefsel in het oog, verzamelt informatie met een neuronaal circuit dat voornamelijk bestaat uit fotoreceptoren, bipolaire cellen en retinale ganglioncellen (RGC’s), de projectieneuronen van het netvlies. RGC’s hebben lange axonen die hun weg vinden over het binnenoppervlak van het netvlies naar de oogzenuwkop, waar ze fasciculeren en samen door de hersenen reizen, uiteindelijk eindigend in het visuele verwerkingscentrum in de dorsale middenhersenen. Deze structuur wordt het optische tectum genoemd bij vissen en andere niet-zoogdiergewervelde dieren en is homoloog aan de superieure colliculus bij zoogdieren3.
Het optische tectum is een bilateraal symmetrische meerlagige structuur in de dorsale middenhersenen. Bij zebravissen en de meeste andere vissen ontvangt elke kwab van het optische tectum alleen visuele input van het contralaterale oog, zodat de linker oogzenuw eindigt in de rechter tectale kwab en de rechter oogzenuw eindigt in de linker tectalkwab4 (figuur 1). Net als zijn tegenhanger bij zoogdieren, de superieure colliculus, integreert het optische tectum visuele informatie met andere sensorische inputs, waaronder auditie en somatosensatie, waarbij verschuivingen in visuele aandacht en oogbewegingen zoals saccades 1,5,6 worden geregeld. In tegenstelling tot de superieure colliculus van zoogdieren genereert het optische tectum echter voortdurend nieuwe neuronen en glia uit een gespecialiseerde stamcelniche in de buurt van de mediale en caudale randen van de tectale kwabben die de tectale proliferatiezone7 worden genoemd. Het behoud van proliferatieve voorlopercellen in het optische tectum en andere regio’s van het centrale zenuwstelsel draagt gedeeltelijk bij aan de opmerkelijke regeneratieve capaciteit die is gedocumenteerd bij zebravissen8.
Eerder onderzoek naar de hersenen van blinde of eenogige vissen onthulde dat de grootte van het optische tectum recht evenredig is met de hoeveelheid retinale innervatie die het ontvangt 9,10,11. Bij volwassen grotvissen, waarvan de ogen degenereren in vroege embryogenese, is het optische tectum merkbaar kleiner dan dat van nauw verwante, waargenomen oppervlaktevissen9. Cave fish eye degeneratie kan worden geblokkeerd door de endogene lens te vervangen door een lens van een oppervlaktevis tijdens embryogenese. Wanneer deze eenogige grotvissen volwassen worden gemaakt, bevat de geïnnerveerde tectale kwab ongeveer 10% meer cellen dan de niet-geïnnerveerde tectale kwab9. Evenzo was bij larvale killivissen die werden geïncubeerd met chemische behandelingen om ogen van verschillende groottes binnen hetzelfde individu te genereren, de kant van het tectum met meer innervatie groter en bevatte meer neuronen10. Bewijs van oogzenuwverpletteringsexperimenten bij volwassen goudvissen geeft aan dat innervatie proliferatie bevordert, waarbij de proliferatie van tectale cellen afneemt wanneer innervatie werd verstoord11.
Om deze klassieke studies te bevestigen en uit te breiden, bieden verschillende recente rapporten gegevens die suggereren dat proliferatie in reactie op innervatie, althans gedeeltelijk, wordt gemoduleerd door de BDNF-TrkB-route12,13. Veel open vragen over de groei en ontwikkeling van optische tectum blijven bestaan, waaronder hoe een zich ontwikkelend sensorisch systeem omgaat met letsel en axondegeneratie, welke cellulaire en moleculaire signalen retinale input mogelijk maken om de groei van het optische tectum te reguleren, wanneer deze mechanismen actief worden, en of innervatie-gekoppelde proliferatie en differentiatie het netvlies en zijn doelweefsel in staat stellen om groeisnelheden te coördineren en nauwkeurige retinotopische mapping te garanderen. Bovendien zijn er veel grotere vragen over activiteitsafhankelijke ontwikkeling die kunnen worden aangepakt door het visuele systeem van zebravissen te ondervragen met chirurgische benaderingen zoals hieronder beschreven.
Om de cellulaire en moleculaire mechanismen te onderzoeken waarmee neurale activiteit, met name uit visuele input, de overleving en proliferatie van cellen verandert, vergelijkt de beschreven benadering direct geïnnerveerde en gedenervateerde tectale kwabben (figuur 1) binnen individuele zebravislarven. Deze methode maakt het mogelijk om RGC axondegeneratie in het optische tectum te documenteren en te bevestigen dat het aantal mitotische cellen correleert met innervatie.
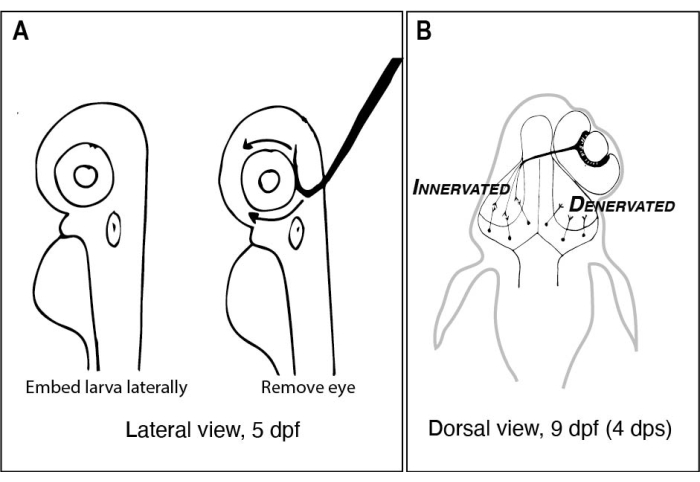
Figuur 1: Schetsen van zebravislarven voor en na eenzijdige oogverwijdering. (A) Tekening van 5 dpflarven gezien onder een ontleedmicroscoop. Elke larve is ingebed in agarose met een laag smeltpunt en zijdelings georiënteerd voordat een wolfraamnaald met een scherpe, gehaakte punt wordt gebruikt om het oog naar boven te scheppen (linkeroog in dit voorbeeld). (B) Tekening van het dorsale beeld van een larve van 9 dpf als gevolg van de in A afgebeelde operatie. Slechts drie sterk geschematiseerde RGC-axonen uit het rechteroog worden getoond die zich defasciculeren en verbinden met neuronen in de linker tectalkwab. Afkortingen: dpf = dagen na de bevruchting; dps = dagen na de operatie; RGC = retinale ganglioncellen. Klik hier om een grotere versie van deze figuur te bekijken.
