Le but de cette méthode est d’étudier comment l’entrée rétinienne influence la croissance et le développement du tectum optique, le centre de traitement visuel dans le cerveau du poisson-zèbre. En enlevant un œil, puis en comparant les deux côtés du tectum optique, les changements tectaux au sein du même spécimen peuvent être observés et normalisés, ce qui permet une comparaison entre plusieurs spécimens. Les approches moléculaires modernes combinées à cette technique permettront de mieux comprendre les mécanismes sous-jacents à la croissance et au développement du système visuel, ainsi qu’à la dégénérescence et à la régénération axonales.
Les systèmes sensoriels – visuels, auditifs et somatosensoriels – recueillent des informations à partir d’organes externes et transmettent ces informations au système nerveux central, générant des « cartes » du monde extérieur à travers le mésencéphale 1,2. La vision est la modalité sensorielle dominante pour presque tous les vertébrés, y compris de nombreux poissons. La rétine, le tissu neural de l’œil, recueille des informations avec un circuit neuronal composé principalement de photorécepteurs, de cellules bipolaires et de cellules ganglionnaires rétiniennes (CGR), les neurones de projection de la rétine. Les RGC ont de longs axones qui se frayent un chemin à travers la surface interne de la rétine jusqu’à la tête du nerf optique, où ils fasciculent et voyagent ensemble à travers le cerveau, se terminant finalement dans le centre de traitement visuel dans le mésencéphale dorsal. Cette structure est appelée tectum optique chez les poissons et autres vertébrés non mammifères et est homologue au colliculus supérieur chez les mammifères3.
Le tectum optique est une structure multicouche à symétrie bilatérale dans le mésencéphale dorsal. Chez le poisson-zèbre et la plupart des autres poissons, chaque lobe du tectum optique reçoit une entrée visuelle provenant uniquement de l’œil controlatéral, de sorte que le nerf optique gauche se termine dans le lobe tectal droit et le nerf optique droit se termine dans le lobe tectal gauche4 (Figure 1). Comme son homologue mammifère, le colliculus supérieur, le tectum optique intègre l’information visuelle à d’autres entrées sensorielles, y compris l’audition et la somatosensation, contrôlant les changements dans l’attention visuelle et les mouvements oculaires tels que les saccades 1,5,6. Cependant, contrairement au colliculus supérieur des mammifères, le tectum optique génère continuellement de nouveaux neurones et glies à partir d’une niche de cellules souches spécialisée près des bords médial et caudal des lobes tectaux appelée zone de prolifération tectale7. Le maintien des progéniteurs prolifératifs dans le tectum optique et d’autres régions du système nerveux central contribue, en partie, à la remarquable capacité de régénération documentée chez le poisson-zèbre8.
Des travaux antérieurs examinant le cerveau de poissons aveugles ou borgnes ont révélé que la taille du tectum optique est directement proportionnelle à la quantité d’innervation rétinienne qu’il reçoit 9,10,11. Chez les poissons des cavernes adultes, dont les yeux dégénèrent au début de l’embryogenèse, le tectum optique est sensiblement plus petit que celui des poissons de surface observésétroitement apparentés 9. La dégénérescence de l’œil de poisson des cavernes peut être bloquée en remplaçant la lentille endogène par une lentille d’un poisson de surface pendant l’embryogenèse. Lorsque ces poissons des cavernes borgnes sont élevés à l’âge adulte, le lobe tectal innervé contient environ 10% plus de cellules que le lobe tectal non innervé9. De même, chez les larves de killifish qui ont été incubées avec des traitements chimiques pour générer des yeux de différentes tailles au sein d’un même individu, le côté du tectum avec plus d’innervation était plus grand et contenait plus de neurones10. Les preuves provenant d’expériences d’écrasement du nerf optique chez des poissons rouges adultes indiquent que l’innervation favorise la prolifération, la prolifération des cellules tectales diminuant lorsque l’innervation est perturbée11.
Confirmant et étendant ces études classiques, plusieurs rapports récents fournissent des données suggérant que la prolifération en réponse à l’innervation est modulée, au moins en partie, par la voie BDNF-TrkB12,13. De nombreuses questions ouvertes sur la croissance et le développement du tectum optique demeurent, notamment comment un système sensoriel en développement fait face aux blessures et à la dégénérescence axonale, quels signaux cellulaires et moléculaires permettent à l’entrée rétinienne de réguler la croissance du tectum optique, quand ces mécanismes deviennent actifs, et si la prolifération et la différenciation liées à l’innervation permettent à la rétine et à son tissu cible de coordonner les taux de croissance et d’assurer une cartographie rétinotopique précise. En outre, il existe des questions beaucoup plus vastes sur le développement dépendant de l’activité qui peuvent être abordées en interrogeant le système visuel du poisson-zèbre avec des approches chirurgicales telles que celle décrite ci-dessous.
Pour étudier les mécanismes cellulaires et moléculaires par lesquels l’activité neuronale, en particulier à partir de l’entrée visuelle, modifie la survie et la prolifération cellulaires, l’approche décrite compare directement les lobes tectaux innervés et dénervés (Figure 1) chez les larves individuelles de poisson zèbre. Cette méthode permet de documenter la dégénérescence des axones RGC dans le tectum optique et de confirmer que le nombre de cellules mitotiques est en corrélation avec l’innervation.
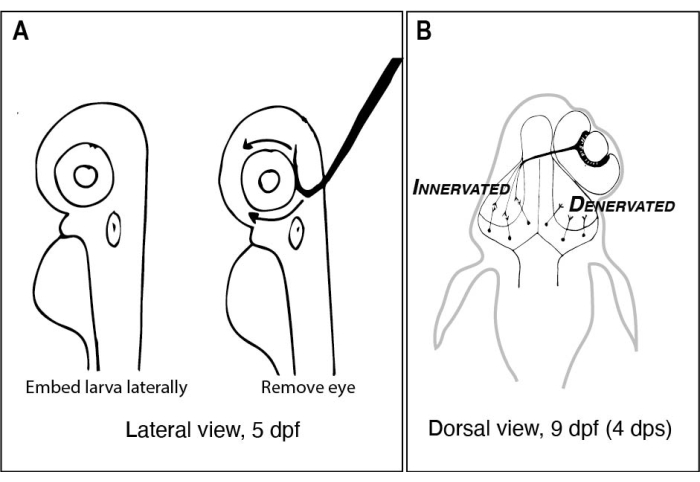
Figure 1: Croquis de larves de poisson zèbre avant et après ablation unilatérale des yeux. (A) Dessin de 5 larves de dpf vues au microscope à dissection. Chaque larve est incrustée dans l’agarose à point de fusion bas et orientée latéralement avant qu’une aiguille en tungstène avec une pointe pointue et crochue ne soit utilisée pour arracher l’œil tourné vers le haut (œil gauche dans cet exemple). (B) Dessin de la vue dorsale d’une larve de 9 dpf résultant de la chirurgie décrite dans A. Seuls trois axones RGC hautement schématisés de l’œil droit sont montrés en train de défasciculer et de se connecter aux neurones du lobe tectal gauche. Abréviations : dpf = jours après la fécondation ; dps = jours après la chirurgie; RGC = cellules ganglionnaires rétiniennes. Veuillez cliquer ici pour voir une version agrandie de cette figure.
