Målet med denne metoden er å undersøke hvordan retinal input påvirker veksten og utviklingen av det optiske tectum, det visuelle prosesseringssenteret i sebrafiskhjernen. Ved å fjerne ett øye og deretter sammenligne de to sidene av det optiske tectumet, kan tektalendringer i samme prøve observeres og normaliseres, noe som muliggjør sammenligning på tvers av flere prøver. Moderne molekylære tilnærminger kombinert med denne teknikken vil gi innsikt i mekanismene som ligger til grunn for visuell systemvekst og utvikling, samt axonal degenerasjon og regenerering.
Sensoriske systemer – visuelle, auditive og somatosensoriske – samler informasjon fra eksterne organer og videresender den informasjonen til sentralnervesystemet, og genererer “kart” over den ytre verden over midbrain 1,2. Visjon er den dominerende sensoriske modaliteten for nesten alle vertebrater, inkludert mange fisk. Netthinnen, nevralt vev i øyet, samler informasjon med en nevronal krets som hovedsakelig består av fotoreseptorer, bipolare celler og netthinne ganglion celler (RGCs), projeksjon nevroner av netthinnen. RGCer har lange axoner som finner veien over den indre overflaten av netthinnen til synsnerven hodet, hvor de fasciculate og reise sammen gjennom hjernen, til slutt avslutte i visuell behandling senter i dorsal midbrain. Denne strukturen kalles det optiske tectum i fisk og andre ikke-pattedyr vertebrater og er homolog til den overlegne colliculus hos pattedyr3.
Det optiske tectum er en bilateralt symmetrisk flerlagsstruktur i dorsal midbrain. I sebrafisk og de fleste andre fisk mottar hver lobe av det optiske tectum visuelle innspill utelukkende fra det kontralaterale øyet, slik at venstre optisk nerve slutter i høyre tectal lobe og høyre optisk nerve slutter i venstre tectal lobe4 (figur 1). Som sin pattedyr motpart, den overlegne colliculus, integrerer det optiske tectum visuell informasjon med andre sensoriske innganger, inkludert audition og somatosensasjon, kontrollerende skift i visuell oppmerksomhet og øyebevegelser som saccades 1,5,6. Men i motsetning til pattedyrets overlegne colliculus, genererer det optiske tectum kontinuerlig nye nevroner og glia fra en spesialisert stamcellenisje nær de mediale og kaudale kantene på tektal lobes kalt tectal proliferation zone7. Vedlikehold av proliferative forfedre i det optiske tectum og andre regioner i sentralnervesystemet bidrar delvis til den bemerkelsesverdige regenerative kapasiteten dokumentert i sebrafisk8.
Tidligere arbeid med å undersøke hjernen til blinde eller enøyde fisk avslørte at optisk tectumstørrelse er direkte proporsjonal med mengden retinal innervering den mottar 9,10,11. I voksenhulefisk, hvis øyne degener i tidlig embryogenese, er det optiske tectum merkbart mindre enn for nært beslektet, observert overflatefisk9. Cave fish eye degenerasjon kan blokkeres ved å erstatte den endogene linsen med en linse fra en overflatefisk under embryogenese. Når disse enøyde hulefiskene blir oppdrettet til voksen alder, inneholder den innerverte tectal lobe omtrent 10% flere celler enn den ikke-innerverte tectal lobe9. På samme måte, i larval killifish som ble inkubert med kjemiske behandlinger for å generere øyne av forskjellige størrelser i samme individ, var siden av tectum med mer innervasjon større og inneholdt flere nevroner10. Bevis fra optisk nerveknusing eksperimenter i voksen gullfisk indikerer at innervasjon fremmer spredning, med tectal celleproliferasjon avtagende når innervasjon ble forstyrret11.
Ved å bekrefte og utvide disse klassiske studiene, gir flere nylige rapporter data som tyder på at spredning som svar på innervering er modulert, i hvert fall delvis, av BDNF-TrkB-banen12,13. Mange åpne spørsmål om optisk tectum vekst og utvikling gjenstår, inkludert hvordan et utviklende sensorisk system håndterer skade og axon degenerasjon, hvilke cellulære og molekylære signaler som muliggjør retinal input for å regulere optisk tectum vekst, når disse mekanismene blir aktive, og om innervasjon-koblet spredning og differensiering gjør det mulig for netthinnen og målvevet å koordinere vekstrater og sikre nøyaktig retinotopisk kartlegging. I tillegg er det mye større spørsmål om aktivitetsavhengig utvikling som kan løses ved å forhøre sebrafiskens visuelle system med kirurgiske tilnærminger som den som er beskrevet nedenfor.
For å undersøke de cellulære og molekylære mekanismene som nevral aktivitet, spesielt fra visuell inngang, endrer celleoverlevelse og spredning, sammenligner den beskrevne tilnærmingen direkte innerverte og denerverte tektallober (figur 1) innenfor individuelle sebrafisk larver. Denne metoden tillater dokumentasjon av RGC axon degenerasjon i det optiske tectum og bekreftelse på at antall mititotiske celler korrelerer med innervasjon.
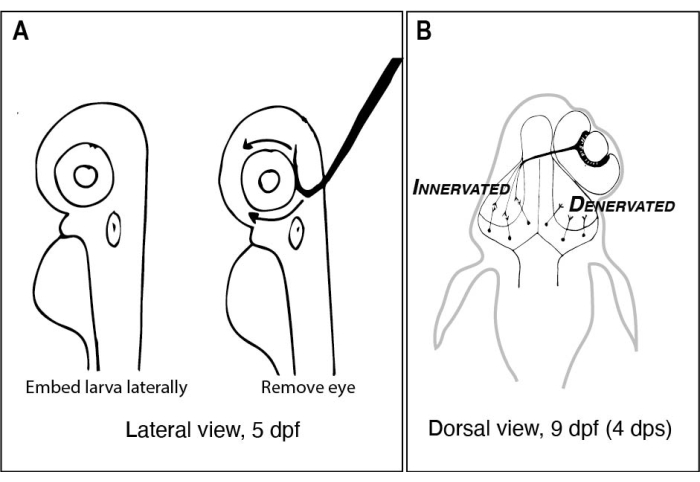
Figur 1: Skisser av sebrafisk larver før og etter ensidig øyefjerning. (A) Tegning av 5 dpf larver sett under et dissekerende mikroskop. Hver larve er innebygd i lavsmeltende punkt agarose og orientert lateralt før en wolfram nål med en skarp, hektet spiss brukes til å øse ut øyet vendt opp (venstre øye i dette eksemplet). (B) Tegning av dorsal utsikt over en 9 dpf larve som følge av operasjonen avbildet i A. Bare tre høyt skjemaerte RGC-axoner fra høyre øye vises defasciculating og tilkobling med nevroner i venstre tectal lobe. Forkortelser: dpf = dager etter befruktning; dps = dager etter operasjonen; RGC = retinal ganglion celler. Klikk her for å se en større versjon av denne figuren.
