O objetivo deste método é investigar como a entrada da retina influencia o crescimento e o desenvolvimento do tectum óptico, o centro de processamento visual no cérebro de zebrafish. Removendo um olho e, em seguida, comparando os dois lados do tectum óptico, alterações tectal dentro do mesmo espécime podem ser observadas e normalizadas, permitindo a comparação entre vários espécimes. Abordagens moleculares modernas combinadas com esta técnica produzirão insights sobre os mecanismos subjacentes ao crescimento e desenvolvimento do sistema visual, bem como a degeneração e regeneração axonal.
Sistemas sensoriais – visuais, auditivos e somatosensoriais – coletam informações de órgãos externos e retransmitem essas informações para o sistema nervoso central, gerando “mapas” do mundo externo em todo o cérebro médio 1,2. A visão é a modalidade sensorial dominante para quase todos os vertebrados, incluindo muitos peixes. A retina, o tecido neural no olho, reúne informações com um circuito neuronal composto principalmente por fotorreceptores, células bipolares e células gânglios da retina (RGCs), os neurônios de projeção da retina. Os RGCs têm longos axônios que encontram seu caminho através da superfície interna da retina até a cabeça do nervo óptico, onde eles fasciculam e viajam juntos através do cérebro, terminando no centro de processamento visual no cérebro dorsal. Esta estrutura é chamada de tectum óptico em peixes e outros vertebrados não-mamíferos e é homóloga ao colegiado superior em mamíferos3.
O tectum óptico é uma estrutura multicamadas bilateralmente simétrica no cérebro médio dorsal. Em zebrafish e na maioria dos outros peixes, cada lóbulo do tectum óptico recebe entrada visual apenas do olho contralateral, de modo que o nervo óptico esquerdo termina no lobo tectal direito e o nervo óptico direito termina no lobo tectal esquerdo4 (Figura 1). Assim como sua contraparte mamífera, o collículo superior, o tectum óptico integra informações visuais com outras entradas sensoriais, incluindo audição e somatosensação, controlando mudanças na atenção visual e movimentos oculares, como saccades 1,5,6. No entanto, ao contrário do colículo superior mamífero, o tectum óptico gera continuamente novos neurônios e glia a partir de um nicho especializado de células-tronco perto das bordas medial e caudal dos lobos tectais chamado zona de proliferação tectal7. A manutenção de progenitores proliferativos no tecto óptico e em outras regiões do sistema nervoso central contribui, em parte, para a notável capacidade regenerativa documentada no zebrafish8.
Trabalhos anteriores examinando os cérebros de peixes cegos ou caolho revelaram que o tamanho do tectum óptico é diretamente proporcional à quantidade de inervação da retina que recebe 9,10,11. Em peixes de cavernas adultos, cujos olhos degeneram em embriogênese precoce, o tectum óptico é visivelmente menor do que o de peixes de superfície9 intimamente relacionados. A degeneração dos olhos dos peixes das cavernas pode ser bloqueada substituindo a lente endógena por uma lente de um peixe superficial durante a embriogênese. Quando esses peixes de caverna de um olho só são criados até a idade adulta, o lobo tectal inervatado contém aproximadamente 10% mais células do que o lobo tectal não inervado9. Da mesma forma, em killifish larval que foram incubados com tratamentos químicos para gerar olhos de diferentes tamanhos dentro do mesmo indivíduo, o lado do tectum com mais inervação foi maior e continha mais neurônios10. Evidências de experimentos de esmagamento de nervo óptico em peixes-dourados adultos indicam que a inervação promove a proliferação, com a proliferação de células tectais diminuindo quando a inervação foi interrompida11.
Confirmando e ampliando esses estudos clássicos, vários relatórios recentes fornecem dados sugerindo que a proliferação em resposta à inervação é modulada, pelo menos em parte, pela via BDNF-TrkB12,13. Muitas questões abertas sobre o crescimento e desenvolvimento do tectum óptico permanecem, incluindo como um sistema sensorial em desenvolvimento lida com lesões e degeneração do axônio, que os sinais celulares e moleculares permitem a entrada da retina para regular o crescimento do tectum óptico, quando esses mecanismos se tornam ativos, e se a proliferação e diferenciação ligadas à invasão permitem que a retina e seu tecido alvo coordenem as taxas de crescimento e garantam um mapeamento retinotópico preciso. Além disso, há questões muito maiores sobre o desenvolvimento dependente da atividade que podem ser abordadas interrogando o sistema visual de zebrafish com abordagens cirúrgicas como a descrita abaixo.
Para investigar os mecanismos celulares e moleculares pelos quais a atividade neural, especificamente a partir da entrada visual, altera a sobrevivência e a proliferação celular, a abordagem descrita compara diretamente os lobos tectal invatados e denervados (Figura 1) dentro de larvas individuais de zebrafish. Este método permite a documentação da degeneração do axônio RGC no tectum óptico e a confirmação de que o número de células mitóticas se correlaciona com a inervação.
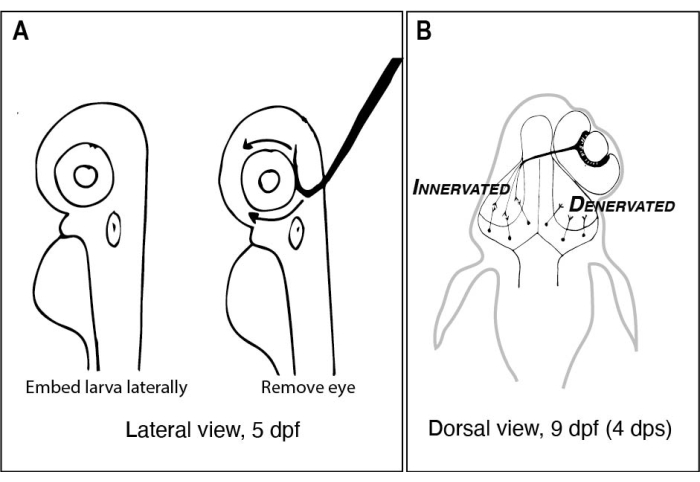
Figura 1: Esboços de larvas de zebrafish antes e depois da remoção unilateral dos olhos. (A) Desenho de 5 larvas dpf vistas sob um microscópio dissecador. Cada larva é embutida em agarose de ponto de derretimento baixo e orientada lateralmente antes que uma agulha de tungstênio com uma ponta afiada e ligada seja usada para colher o olho voltado para cima (olho esquerdo neste exemplo). (B) Desenho da visão dorsal de uma larva de 9 dpf resultante da cirurgia retratada em A. Apenas três axônios RGC altamente esquematizados do olho direito são mostrados desfigurando e conectando-se com neurônios no lobo tectal esquerdo. Abreviaturas: dpf = dias pós fertilização; dps = dias após a cirurgia; RGC = células gânglios de retina. Clique aqui para ver uma versão maior desta figura.
