De bodem is een uitzonderlijk diverse omgeving met een overvloed aan micro-organismen die een belangrijke rol spelen voor koolstof- en fosforcycli 1,2. Filamenteuze schimmels zijn een belangrijk onderdeel van talrijke ecosystemen als afbrekers van organische en anorganische materie en kunnen de voeding van planten verbeteren door de vorming van symbiotische relaties 3,4. In de bodem interageren schimmels dynamisch met een veelheid aan microben zoals andere schimmels5, bacteriën6, virussen7 en nematoden8. Deze interacties hebben aanzienlijke gevolgen voor de bodem- en plantgezondheid. Maar als gevolg van een gebrek aan geschikte experimentele systemen die in staat zijn om interagerende micro-organismen met hoge resolutie in beeld te brengen, blijven velen ongedefinieerd.
Onderzoeken met betrekking tot bacteriële-schimmelinteracties (BFI’s) en schimmel-schimmelinteracties (FFI’s) hebben waardevolle toepassingen op verschillende gebieden, waaronder antimicrobiële stoffen in de geneeskunde en biologische bestrijders in de landbouw. De schimmel Coprinopsis cinerea produceert bijvoorbeeld het peptide copsine, waarvan is aangetoond dat het antibacteriële activiteit vertoont tegen de menselijke ziekteverwekker Listeria monocytogenes9. Evenzo wordt de van schimmel afgeleide verbinding, griseofulvine, veel gebruikt als behandeling voor menselijke schimmelinfecties en is bovendien in staat om de groei van de plantpathogene schimmel Alternaria solani10,11 te remmen. Verschillende stammen van de bodembewonende bacterie Bacillus subtilis zijn ook effectieve biocontrolemiddelen gebleken van de schimmelplantpathogeen Rhizoctonia solani12,13. Niettemin, als gevolg van beperkingen in verband met traditionele methodologieën, worden BFI’s en FFI’s slecht begrepen op het niveau van enkele cellen.
Conventionele studies onderzoeken meestal BFI’s en FFI’s op macroschaal met behulp van agarplaten met twee of meer soorten in confrontatie. Hun interactie wordt beoordeeld door het meten van groeisnelheden en metabolietproductie van de confronterende soort 14,15,16; deze methodologie wordt echter alleen opgelost tot op kolonieniveau. Om interacties op cellulair niveau te bestuderen, kunnen bacteriële en schimmelinoculanten worden gekweekt op glazen microscoopglaasjes bedekt met agar die vervolgens worden afgebeeld onder een microscoop17. Niettemin kan het moeilijk zijn om een enkele hypha te volgen met behulp van microscoopglaasjes vanwege een gebrek aan opsluiting, wat betekent dat time-lapse-beelden moeilijker te verkrijgen zijn. Verder is de mogelijkheid om andere micro-organismen ruimtelijk op te sluiten binnen gedefinieerde gebieden van het schimmelmycelium of gedefinieerde chemische omgevingen te creëren die bijvoorbeeld kunnen worden verstoord, niet mogelijk in dergelijke opstellingen. De “black box” aard van de bodem draagt ook bij aan de complexiteit van het bestuderen van schimmel-microbiële interacties op het niveau van enkele cellen18. Door interagerende soorten te observeren, weg van de ongelooflijke diversiteit van het bodemmicrobioom, kan de exacte manier waarop individuele leden met elkaar omgaan worden beoordeeld. Er is dus een voortdurende behoefte aan veelzijdige platforms die hoge-resolutie, single-cell imaging van BFI’s en FFI’s mogelijk maken.
Microfluïdische technologieën, zogenaamde lab-on-a-chip-systemen, bieden een ideaal platform voor de studie van BFI’s en FFI’s op het niveau van enkele cellen. Het gebied van microfluïdica, afkomstig van technologieën die zijn ontwikkeld voor chemische analyse en micro-elektronica, is overgenomen door de biologische wetenschappen19. Microfluïdische technologieën reguleren kleine hoeveelheden vloeistoffen binnen een op maat gemaakt netwerk van geminiaturiseerde kanalen, met ten minste één dimensie op micrometerschaal, en hun gebruik in biologisch onderzoek breidt zich uit20. In het bijzonder zijn microfluïdische apparaten ontwikkeld om de groei van filamenteuze schimmels 21,22,23,24,25,26,27,28,29,30 te onderzoeken. Een voordeel van het gebruik van deze technologie is dat de opsluiting van schimmeldraden en de verdeling van voedingsstoffen in microkanalen meer lijkt op de structuur van het bodemmilieu dan conventionele agarmethoden31. Onlangs zijn microfluïdische platforms gebruikt om interacties tussen menselijke neutrofielen en schimmelpathogenen32, bacteriën en plantenwortels33, evenals schimmels en nematoden34,35 te onderzoeken.
Een van de vele voordelen van het gebruik van microfluïdica voor het bestuderen van microbiële interacties omvat de specifieke controle van de microkanaalomgeving. Laminaire stromingsregimes kunnen bijvoorbeeld worden gebruikt om gedefinieerde concentratiegradiënten te genereren, wat vooral handig is bij het onderzoeken van bacteriële chemotaxis36. Een ander voordeel is dat de transparante aard van poly (dimethylsiloxaan) (PDMS), een goedkoop, biocompatibel elastomeerpolymeer dat vaak wordt gebruikt bij de vervaardiging van microfluïdische apparaten, beeldvorming met hoge resolutie van afzonderlijke cellen met behulp van brightfield- en fluorescentiemicroscopiemogelijk maakt 37. Evenzo betekent de opsluiting van microben in microkanalen dat time-lapse-experimenten kunnen worden uitgevoerd die afzonderlijke cellen volgen, waardoor individuele cellulaire reacties kunnen worden geregistreerd en gekwantificeerd37. Ten slotte, aangezien microfluïdische apparaten kunnen worden ontworpen om gebruiksvriendelijk te zijn, kunnen ze gemakkelijk worden gebruikt door niet-experts38.
Het bevorderen van de kennis van de interacties tussen bodembewonende micro-organismen is belangrijk voor het verbeteren van duurzame ecosysteembeheerpraktijken die de biodiversiteit in stand houden en om de gevolgen van klimaatverandering voor terrestrische milieus te beperken39. De ontwikkeling van nieuwe microfluïdische hulpmiddelen is dus van fundamenteel belang om het begrip van schimmels en hun interacties op cellulair niveau uit te breiden. Het protocol hier zal zich richten op twee microfluïdische apparaten die zijn geproduceerd voor de studie van BFI’s40 en FFI’s41 zoals weergegeven in figuur 1.
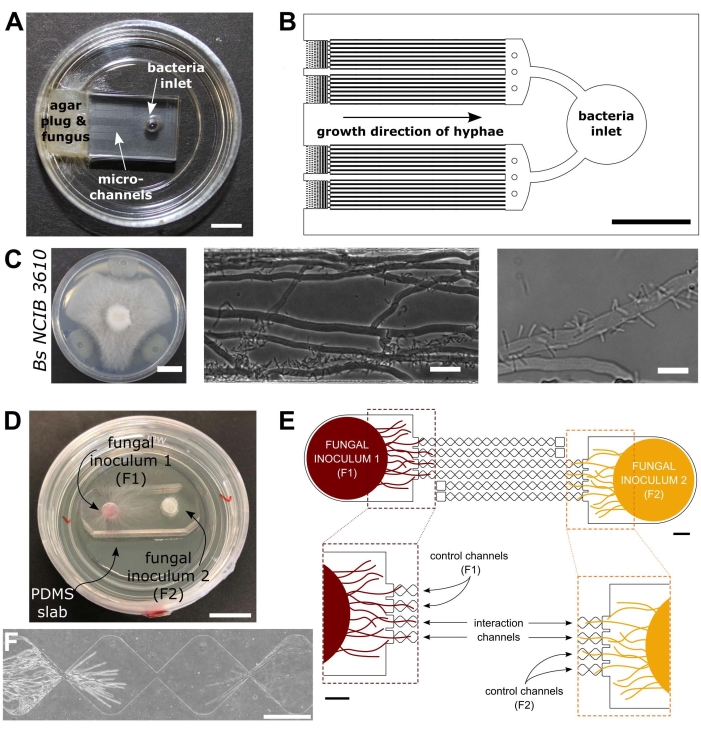
Figuur 1: Visuele en schematische weergave van de bacteriële-schimmelinteractie (BFI) en schimmel-schimmelinteractie (FFI) apparaten. (A) Afbeelding van het BFI-apparaat. Een myceliale plug wordt geplaatst bij de ingang van het ene uiteinde van de microkanalen om hyphale groei in het apparaat mogelijk te maken. De bacteriële inlaat bevindt zich aan de andere kant. Schaalbalk = 5 mm. (B) Schematisch overzicht van het BFI-apparaat, met de positionering van de bacteriële inlaten en de richting van de hyphale groei door de interactiemicrokanalen. De kanalen zijn 10 μm diep, 100 μm breed en 7 mm lang, met in totaal 28 observatiekanalen. (C) Confrontatietest op agarplaat tussen Coprinopsis cinerea en Bacillus subtilis NCIB 3610, schaalbalk = 20 mm (links). Microscopiebeelden die de interactie tonen tussen C. cinerea en B. subtilis NCIB 3610 binnen het microkanaal (midden en rechts), d.w.z. polaire aanhechting van bacteriën aan schimmeldraden. Schaalbalk = 25 μm (midden) en 10 μm (rechts). (D) Afbeelding van het FFI-apparaat dat is verbonden met een petrischaal met glazen bodem, dubbel ingeënt met myceliale pluggen. Schaalbalk = 1 cm. (E) Schematisch overzicht van het FFI-apparaat. Twee schimmelinoculante pluggen worden in de inlaten aan beide uiteinden van het apparaat gebracht, waardoor hyphale exploratie van de microkanalen mogelijk is. Controlekanalen zijn slechts verbonden met één schimmelinlaat en hebben een doodlopend kanaal, waardoor interacties tussen de testschimmels worden voorkomen. Interactiekanalen verbinden beide schimmelinlaten en maken hyphale interacties tussen de proefpersonen in het microkanaal mogelijk. Elk interactiekanaal bestaat uit 18 diamantvormige secties, met een totale lengte van 8,8 mm (490 x 430 μm per diamant), 10 μm diep en met een verbindingsgebied tussen elke diamant van 20 μm. Kanaaltypen worden gedupliceerd, schaalbalken = 1 mm. (F) Interactiezone tussen twee naderende hyphale fronten, groeiend uit tegenovergestelde uiteinden van het onderling verbonden interactiekanaal. Fasecontrastmicroscopiebeeld, schaalbalk = 250 μm. De panelen in deze figuur zijn aangepast van Stanley et al., 2014 (A-C)40 en Gimeno et al., 2021 (D-F)41. Klik hier om een grotere versie van deze figuur te bekijken.
