Jord er et eksepsjonelt mangfoldig miljø som inneholder en overflod av mikroorganismer som er medvirkende til karbon- og fosforsykluser 1,2. Filamentøse sopp er en viktig komponent i mange økosystemer som nedbrytere av organisk og uorganisk materiale og kan forbedre ernæringen av planter gjennom dannelsen av symbiotiske forhold 3,4. Innenfor jord samhandler sopp dynamisk med en rekke mikrober som andre sopp5, bakterier6, virus7 og nematoder8. Disse interaksjonene har betydelige konsekvenser for jord- og plantehelsen. Likevel, på grunn av mangel på passende eksperimentelle systemer som er i stand til å avbilde interagerende mikroorganismer med høy oppløsning, forblir mange udefinerte.
Undersøkelser om bakteriell-sopp interaksjoner (BFF) og sopp-sopp interaksjoner (FFIer) har verdifulle anvendelser på en rekke felt, inkludert antimikrobielle midler i medisin og biologiske kontrollmidler i landbruket. For eksempel produserer sopp coprinopsis cinerea peptid copsin, som har vist seg å vise antibakteriell aktivitet mot det menneskelige patogenet Listeria monocytogenes9. På samme måte er den soppavledede forbindelsen, griseofulvin, mye brukt som behandling for menneskelige soppinfeksjoner og er i tillegg i stand til å hemme veksten av planten patogen sopp Alternaria solani10,11. Flere stammer av jordboende bakterie Bacillus subtilis har også vist seg å være effektive biokontrollmidler av soppplantepatogenet Rhizoctonia solani12,13. Likevel, på grunn av begrensninger knyttet til tradisjonelle metoder, er BBFIer og FFIer dårlig forstått på nivået av enkeltceller.
Konvensjonelle studier utforsker vanligvis BBFIer og FFIer på makroskalaen ved hjelp av agarplater med to eller flere arter i konfrontasjon. Deres interaksjon vurderes ved å måle vekstrater og metabolittproduksjon av de konfronterende artene 14,15,16; Denne metodikken løses imidlertid bare til koloninivå. For å studere interaksjoner på cellenivå, kan bakterielle og soppinokulanter dyrkes på glassmikroskopsklier belagt med agar som deretter avbildes under et mikroskop17. Likevel kan det være vanskelig å følge en enkelt hypha ved hjelp av mikroskoplysbilder på grunn av mangel på innesperring, noe som betyr at tidsforløpbilder er vanskeligere å få tak i. Videre er muligheten til å romlig begrense andre mikroorganismer innenfor definerte regioner i soppmiceliet eller skape definerte kjemiske miljøer som for eksempel kan bli forstyrret, ikke mulig i slike oppsett. Jordens “black box” natur legger også til kompleksiteten ved å studere sopp-mikrobielle interaksjoner på nivået av enkeltceller18. Ved å observere interagerende arter vekk fra det utrolige mangfoldet i jordmikrobiomet, kan den nøyaktige måten individuelle medlemmer samhandler på vurderes. Dermed er det et fortsatt behov for allsidige plattformer som muliggjør høyoppløselig, encellet bildebehandling av BFFIer og FFIer.
Mikrofluidisk teknologi, såkalte lab-on-a-chip-systemer, gir en ideell plattform for studiet av BFF-er og FFIer på nivået av enkeltceller. Feltet mikrofluidikk, med opprinnelse fra teknologier utviklet for kjemisk analyse og mikroelektronikk, er vedtatt av biovitenskapene19. Mikrofluidiske teknologier regulerer små mengder væsker i et skreddersydd nettverk av miniatyriserte kanaler, har minst en dimensjon på mikrometreskalaen, og bruken i biologisk forskning utvides20. Spesielt er mikrofluidiske enheter utviklet for å undersøke veksten av filamentøse sopp 21,22,23,24,25,26,27,28,29,30. En fordel med å bruke denne teknologien er at innesperring av hyphae og fordelingen av næringsstoffer i mikrokanal ligner mer på jordens struktur enn konvensjonelle agarmetoder31. Nylig har mikrofluidiske plattformer blitt brukt til å undersøke interaksjoner mellom menneskelige nøytrofiler og sopppatogener32, bakterier og planterøtter33, samt sopp og nematoder34,35.
En av de mange fordelene ved å bruke mikrofluidikk for å studere mikrobielle interaksjoner inkluderer den spesifikke kontrollen av mikrokanalmiljøet. For eksempel kan laminære strømningsregimer utnyttes for å generere definerte konsentrasjonsgradienter, noe som er spesielt nyttig når du undersøker bakteriell chemotaxis36. En annen fordel er at den gjennomsiktige naturen til poly (dimetylsiloksan) (PDMS), en billig, biokompatibel elastomerisk polymer som vanligvis brukes til fremstilling av mikrofluidiske enheter, letter høyoppløselig avbildning av enkeltceller ved hjelp av brightfield og fluorescensmikroskopi37. På samme måte betyr innesperring av mikrober i mikrokanal at tidsforløpeksperimenter som sporer enkeltceller, kan utføres, slik at individuelle cellulære responser kan registreres og kvantifiseres37. Til slutt, da mikrofluidiske enheter kan utformes for å være brukervennlige, kan de enkelt brukes av ikke-eksperter38.
Videre kunnskap om samspillet mellom jordboende mikroorganismer er viktig for å forbedre bærekraftige økosystemforvaltningspraksiser som opprettholder biologisk mangfold og for å redusere virkningen av klimaendringer på terrestriske miljøer39. Dermed er utviklingen av nye mikrofluidiske verktøy grunnleggende for å utvide forståelsen av sopp og deres interaksjoner på cellenivå. Protokollen her vil fokusere på to mikrofluidiske enheter produsert for studiet av BFIer40 og FFIer41 som representert i figur 1.
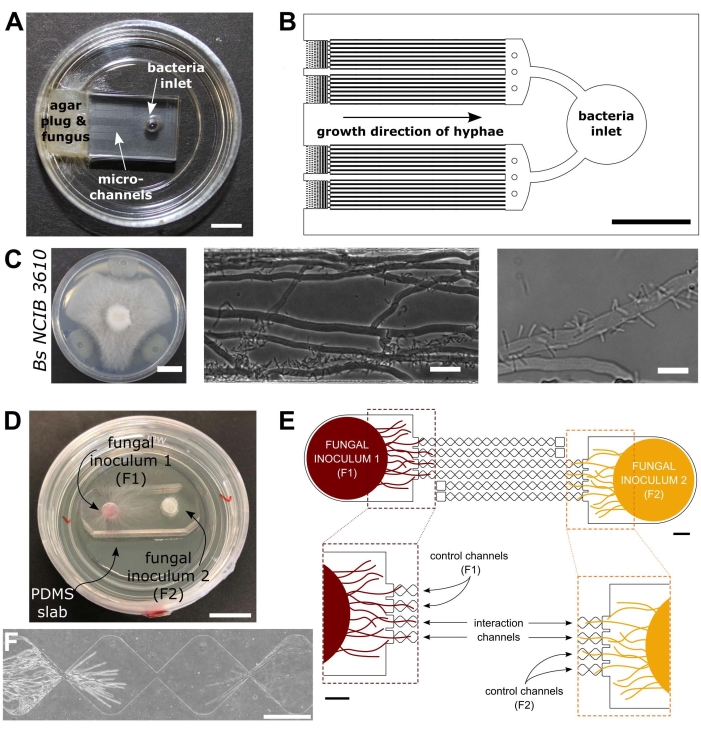
Figur 1: Visuell og skjematisk fremstilling av bakteriell soppinteraksjon (BFI) og sopp-soppinteraksjonsenheter (FFI). (A) Bilde av BFI-enheten. En mycelial plugg er plassert ved inngangen til den ene enden av mikrokanalene for å tillate hyphalvekst i enheten. Bakterieinntaket er i motsatt ende. Skalastang = 5 mm. (B) Skjematisk oversikt over BFI-enheten, som viser plasseringen av bakterieinntakene og retningen av hyphalvekst gjennom interaksjonsmikrokanalene. Kanalene er 10 μm i dybden, 100 μm brede og 7 mm lange, med totalt 28 observasjonskanaler. (C) Konfrontasjonsanalyse på agarplate mellom Coprinopsis sinerea og Bacillus subtilis NCIB 3610, skala bar = 20 mm (venstre). Mikroskopibilder som viser samspillet mellom C. cinerea og B. subtilis NCIB 3610 innenfor mikrokanal (midten og høyre), det vil si polar festing av bakterier til sopphyphae. Skalastang = 25 μm (midten) og 10 μm (høyre). (D) Bilde av FFI-enheten festet til en glassbunnet Petri-tallerken, dobbelt inokulert med mycelialplugger. Skalastang = 1 cm. (E) Skjematisk oversikt over FFI-enheten. To soppinokulantplugger blir introdusert i innløpene i hver ende av enheten, slik at hyphalutforskning av mikrokanalene tillater hyphalutforskning av mikrokanalene. Kontrollkanaler er kun koblet til ett soppinntak og har en blind kanal, noe som forhindrer interaksjoner mellom testsvampene. Interaksjonskanaler forbinder både soppinntak og tillater hyphalinteraksjoner mellom testpersonene i mikrokanal. Hver interaksjonskanal består av 18 diamantformede seksjoner, som måler en total lengde på 8,8 mm (490 x 430 μm per diamant), 10 μm dyp og har en forbindelsesregion mellom hver diamant på 20 μm. Kanaltyper dupliseres, skalastenger = 1 mm. (F) Interaksjonssone mellom to nærliggende hyphalfronter, som vokser fra motsatte ender av den sammenkoblede samhandlingskanalen. Fasekontrastmikroskopibilde, skalalinje = 250 μm. Panelene i denne figuren er modifisert fra Stanley et al., 2014 (A-C)40 og Gimeno et al., 2021 (D-F)41. Klikk her for å se en større versjon av denne figuren.
