Chick embryo survival rates
The embryo survival rate from windowing (day 3 of ED) to ovarian tissue grafting (day 7 of ED) was 79% (33/42). Since the percentage of embryonated day 0 eggs is unknown, supernumerary day 0 eggs from Lohman-selected white Leghorn chickens were ordered to ensure sufficient embryonated eggs would be available for grafting. A total of 23 viable day 7 eggs were used for grafting, one of which perished during the first 24 h, resulting in an overall embryo survival rate of 96% after transplantation (22/23).
Macroscopic aspect of the grafts
After 1 day of grafting, the grafts looked viable in 100% (10/10) of cases and were already at least partially adherent to the CAM, showing a wheel-spoke pattern of blood vessels needed for their vascularization (Figure 2A,B). After 6 days of grafting, all the implants were still adherent (Figure 2C), apart from two that did not attach to the CAM. They took on a necrotic appearance and were excluded from further analysis (Figure 3), resulting in a grafted tissue survival rate of 83% (10/12) after 6 days. All in all, the viability assessment of the human ovarian grafts revealed an overall tissue survival rate of 91% (20/22), irrespective of the day of grafting (Table 2).
Around day 3 post-transplantation, the grafts were found to be covered with a second layer of CAM, and they eventually became encapsulated, leading to even better graft vascularization (Figure 2C). Overall, 80% of the transplants had also penetrated the egg (Figure 2D), in some cases making it hard to retrieve them on day 6.
Microscopic assessment of the grafts
A total of 30 frozen-thawed human ovarian fragments obtained from five different patients were analyzed and fixed in 4% paraformaldehyde on grafting day 0, day 1, or day 6, embedded in paraffin, and serially cut into 5 µm sections for histological evaluation (Figure 4).
In order to assess the ovarian follicle survival rates, follicles were counted in 12 random hematoxylin and eosin-stained sections per time point and per patient. Only morphologically normal follicles with a visible oocyte were taken into consideration for analysis32. Healthy follicles were observed at all time points (Figure 4B–D). The follicle survival rates were calculated by determining the remaining follicle density (number of follicles/mm3) after grafting and normalizing it to the follicle density in non-grafted controls (considered 100%). The follicle densities tended to decrease after transplantation, but not enough to reach statistical significance (p > 0.8) (Table 2). Similarly, the follicle survival rates were maintained during the grafting period, standing at 95% ± 19% on day 1 and 83% ± 27% on day 6 (p > 0.5).
The revascularization process was further investigated by the detection of avian red blood cells in vessels from the xenografted ovarian tissue. Avian erythrocytes are easily discernible since they are nucleated (Figure 4A,C,D). Surprisingly, we were able to observe avian erythrocytes in ovarian vessels in 30% of implants after only 1 day of transplantation and in all the implants by day 6, exhibiting very rapid revascularization (Table 2).

Figure 1: Egg incubator. Please click here to view a larger version of this figure.
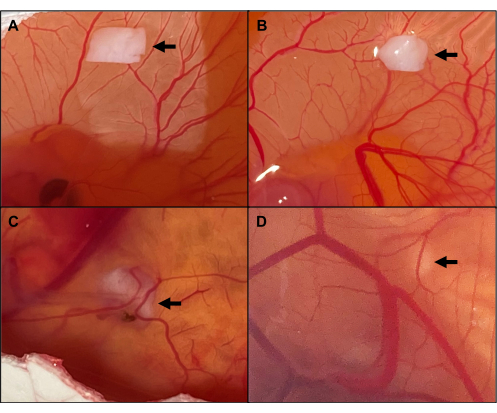
Figure 2: Macroscopic aspect of viable implants. (A) Grafting day 0. (B) Implants on day 1, with the CAM showing a wheel-spoke pattern of blood vessels towards the ovarian tissue. (C) Implants encapsulated by CAM on day 6, leading to even better graft vascularization. (D) Graft penetrating the egg. The arrows point to the grafted ovarian tissue. Please click here to view a larger version of this figure.
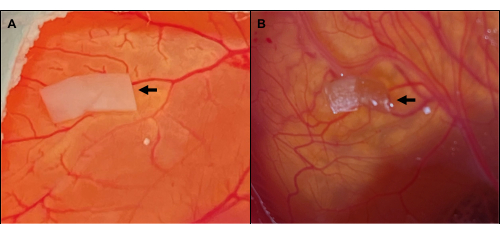
Figure 3: Macroscopic aspect of necrotic implants. (A) Healthy-looking ovarian tissue on day 0 of grafting. (B) Necrotic aspect of the implant 6 days later. The arrows point to the ovarian tissue. Please click here to view a larger version of this figure.
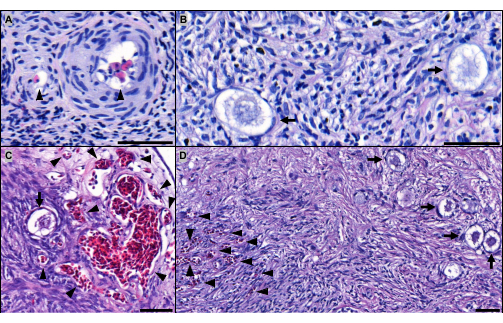
Figure 4: Microscopic aspect of implants. Hematoxylin and eosin-stained sections of implants grafted for (A,B) 1 day and for (C,D) 6 days. The arrowheads point to avian red blood cells perfusing the ovarian vessels after (A) 1 day and (C,D) 6 days of grafting. The arrows indicate healthy-looking primordial follicles in implants grafted for (B) 1 day and (C,D) 6 days. Scale bar: 50 µm. Please click here to view a larger version of this figure.
| Steps | Baths | Time |
| Second fixation | 1 Formalin 10% bath | 2 h |
| Dehydration | 7 methanol baths | 7x 1 h |
| Clarification | 3 toluene baths | 3x 1 h |
| Impregnation | 3 liquid paraffin baths (60°C) | 15 min – 30 min – 30 min |
Table 1: Paraffin embedding steps.
| Non-grafted | Grafting day 1 | Grafting day 6 | P-value | |
| Macroscopic aspect of grafts | ||||
| Tissue survival rate | – | 100% (10/10) | 83% (10/12) | / |
| Microscopic aspect of grafts | ||||
| Follicle density (n/mm3) | 848 ± 272 | 817 ± 370 | 684 ± 236 | p > 0.8 |
| Follicle survival rate | 100% | 95% ± 19% | 83% ± 27% | p > 0.5 |
| Avian red blood cells present in implants | 0% (0/10) | 30% (3/10) | 100% (10/10) | / |
Table 2: Results. Statistical analysis conducted using one-way ANOVA, followed by Sidak correction. Results are expressed as mean ± standard deviation.
