As per the protocol, we have created a bioactive scaffold by dripping BMP-2 into a degradable gelatin sponge under sterile conditions. The scaffold was then implanted into the lower limb muscles of mice to establish in vivo osteo-organoids. After an incubation period of 12 weeks, we conducted macroscopic photography, histological analysis, and flow cytometry analysis on the osteo-organoids (Figure 1A). The gelatin sponge was cut into cubes with dimensions of 5 mm x 5 mm x 5 mm, and after absorbing 30 µL of BMP-2 stock solution (1.0 mg/mL), a bioactive scaffold was obtained. The bioactive scaffold retained its original shape even after freeze-drying (Figure 1B). Scanning electron microscopy (SEM) images depict a porous structure within the bioactive scaffold that promotes cell adhesion and proliferation (Figure 1C). The interconnections among various pores could enhance blood vessel sprouting and invasion, facilitating nutrient transport to support cell growth and aiding in osteo-organoid formation. After 12 weeks of induction in vivo, the implanted osteo-organoids exhibited a deep red color (Figure 1D). H&E-stained image showed that the bioactive scaffold induced the formation of a natural bone marrow-like structure composed of osteocytes and bone marrow cells, further confirming the formation of in vivo osteo-organoids (Figure 1E).
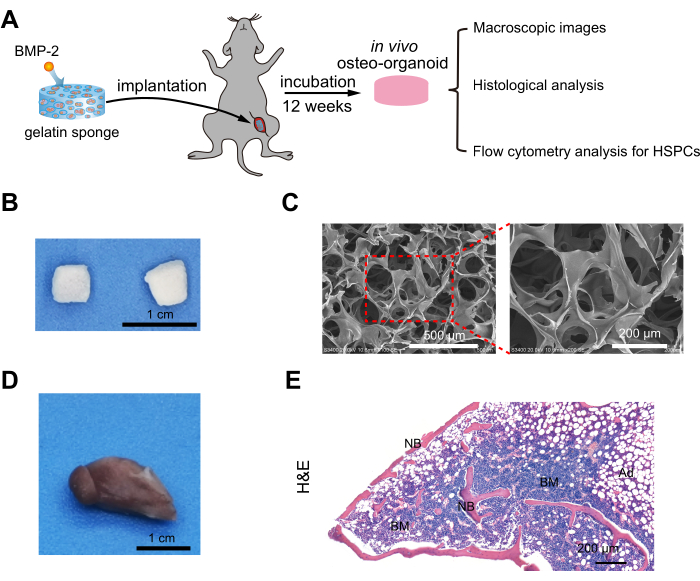
Figure 1: Formation and characterization of in vivo osteo-organoids. (A) Schematic diagram for harvesting and evaluating in vivo osteo-organoids induced by bioactive scaffolds. Freeze-dried gelatin scaffolds loaded with BMP-2 were surgically implanted into the lower limb muscles of mice. The in vivo osteo-organoids were explanted after 12 weeks of incubation for subsequent analysis. (B) Macroscopic images and (C) scanning electron microscope images of the bioactive scaffold. Figure 1A–C are reproduced from Dai et al.11. (D) Macroscopic image and (E) H&E-stained sections of the osteo-organoid 12 weeks after implantation. Figure 1D,E are reproduced from Dai et al.10. n = 4 biological replicates. Bone marrow and new bone are shown. Scale bars = 200 µm (C, right; E), 500 µm (C, left), 1 cm (B,D). Abbreviations: BMP-2 = bone morphogenetic protein-2; H&E = hematoxylin and eosin; BM = bone marrow; NB = new bone. Please click here to view a larger version of this figure.
To assess the long-term therapeutic cell-generating potential of osteo-organoids, we analyzed single-cell suspensions from the osteo-organoids using flow cytometry for quantitative analysis of HSPCs (Supplemental Figure S1). We analyzed the types of HSPC subsets 12 weeks after implantation (Figure 2A). Hematopoietic stem cells (HSCs; Lin−c-kit+Sca-1+CD48−CD150+) and their differentiated hematopoietic progenitor cells (HPCs), including multipotent progenitors (MPPs; Lin−c-kit+Sca-1+CD48−CD150−), common lymphoid progenitors (CLPs; Lin−c-kit–Sca-1–Flt3+IL7Rα+), common myeloid progenitors (CMPs; Lin−c-kit+Sca-1−CD34+FCγR−), granulocyte-monocyte progenitors (GMPs; Lin−c-kit+Sca-1−CD34+FCγR+), and megakaryocyte erythroid progenitors (MEPs; Lin−c-kit+Sca-1−CD34−FCγR−) were all present in the osteo-organoids. We then conducted further quantitative analysis of the cell numbers in different cell populations within the osteo-organoids (Figure 2B). After 12 weeks of implantation, the numbers of HSCs, MMPs, CLPs, CMPs, GMPs, and MEPs in the osteo-organoids were approximately 530, 860, 2750, 80,000, 43,000, and 55,000, respectively. In conclusion, these results supported that osteo-organoids contained an abundant supply of HSPCs for cell therapy.
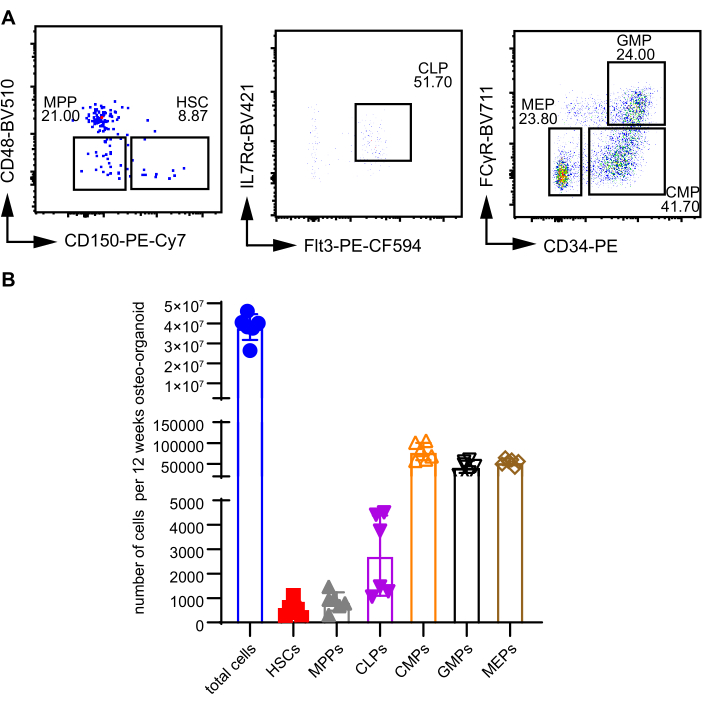
Figure 2: In vivo osteo-organoids contain abundant HSPCs. (A) Representative flow cytometric plots of HSPCs in the osteo-organoids 12 weeks after implantation. (B) Absolute numbers of total cells, Hematopoietic stem cells (Lin−c-kit+Sca-1+CD48−CD150+) and differentiated hematopoietic progenitors: multipotent progenitors (Lin−c-kit+Sca-1+CD48−CD150−), common lymphoid progenitors (Lin−c-kit–Sca-1–Flt3+IL7Rα+), common myeloid progenitors (Lin−c-kit+Sca-1−CD34+FCγR−), granulocyte-monocyte progenitors (Lin−c-kit+Sca-1−CD34+FCγR+), and megakaryocyte erythroid progenitors (Lin−c-kit+Sca-1−CD34−FCγR−). Figure 2A,B was modified from Dai et al.10. n = 6 biological replicates. Data are presented as means ± SD. Statistical differences among groups are identified by one-way ANOVA, followed by Tukey's multiple comparison tests. *P < 0.05, **P < 0.01, and ***P < 0.001. Abbreviations: HSPCs = hematopoietic stem/progenitor cells; HSCs = Hematopoietic stem cells; MPPs = multipotent progenitors; CLPs = common lymphoid progenitors; CMPs = common myeloid progenitors; GMPs = granulocyte-monocyte progenitors; MEPs = megakaryocyte erythroid progenitors; PE = phycoerythrin. Please click here to view a larger version of this figure.
HSCT has proven to be an effective cell therapy for treating radiation-induced immune system injury12,13,14. We transplanted cells derived from in vivo osteo-organoids (incubated for 12 weeks) into sublethally irradiated mice and collected samples from different parts of the mice at indicated time points for subsequent analysis (Figure 3A). Hematological analysis showed that, compared to the native BM-treated group, the WBC count in the 5 Gy osteo-organoid-treated group significantly increased at the 4th week post transplantation, suggesting an enhanced WBC recovery capability of the osteo-organoid-derived cells (Figure 3B). In the second week after transplantation, the RBC count of both the 5 Gy osteo-organoid-treated group and the 5 Gy native BM-treated group returned to normal levels (Figure 3C). At 2 weeks post transplantation, the PLT count was significantly increased in the 5 Gy osteo-organoid-treated group and the 5 Gy native BM-treated group compared to the 5 Gy PBS-treated group (Figure 3D). In conclusion, these results confirmed that cells derived from osteo-organoids exerted a more robust capacity to enhance hematopoietic recovery compared to native BM cells.
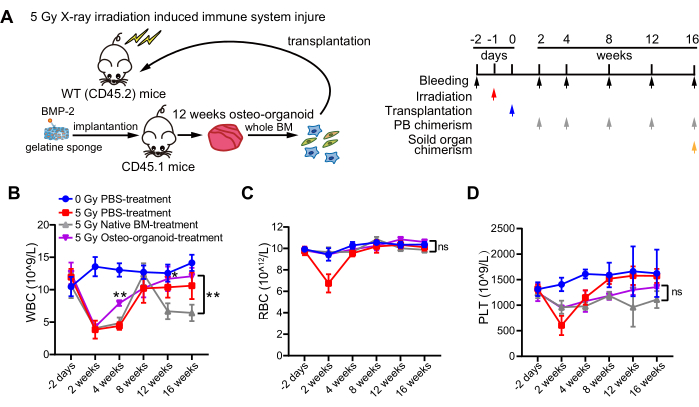
Figure 3: Cell therapy from osteo-organoids for the treatment of sublethally irradiated mice. (A) Experimental scheme of HSCT. The osteo-organoids were generated by implanting bioactive scaffolds into the femur muscle pocket of mice (donors, CD45.1) for 12 weeks. The osteo-organoids were collected and digested to obtain a whole bone marrow cell suspension, which was then transplanted to WT mice (recipients, CD45.2) with immune system injury induced by 5 GY X-ray. peripheral blood was collected at 2, 4, 8, 12, and 16 weeks after transplantation for subsequent analysis of peripheral blood chimerism. Solid organ was collected at 16 weeks after transplantation for subsequent analysis of solid organ chimerism. (B–D) peripheral blood cells from the four groups were analyzed at indicated time points. WT mice in the 0 Gy PBS-treated group did not receive irradiation and received PBS. WT mice in the 5 Gy PBS-treated, 5 Gy native BM-treated, and 5 Gy osteo-organoid-treated groups received 5 Gy irradiation 1 day prior to transplantation and were transplanted with PBS, native BM cells, or osteo-organoid-derived cells, respectively. n = 4 to 5 biological replicates. Figure 3A–D was modified from Dai et al.10. Data are presented as means ± SD. (B–D) Significant differences among groups are identified by two-way ANOVA followed by Bonferroni's post hoc test. **P < 0.01. Abbreviations: HSCT = Hematopoietic stem cell transplantation; BMP-2 = bone morphogenetic protein-2; WT = wild type; PB = peripheral blood; PBS = phosphate-buffered saline; BM = bone marrow; WBC = white blood cell; RBC = red blood cell; PLT = platelet. Please click here to view a larger version of this figure.
To assess the contribution of transplanted cells to the peripheral immune system reconstitution in irradiated mice, we performed quantitative analysis of CD3+ T cells, B220+ B cells, and CD11b+ myeloid cells using flow cytometry. This allowed us to evaluate the chimerism of osteo-organoid-derived and native BM-derived cells in the peripheral blood of the recipients. (Figure 4A and Supplemental Figure S2). We collected peripheral blood samples at 2, 4, 8, 12, and 16 weeks after cell therapy in different groups (Figure 3A). We observed sustained high levels of chimerism in the peripheral blood, with donor-derived CD3+ T cells, B220+ B cells, and CD11b+ myeloid cells consistently exceeding 80% in both the 5 Gy osteo-organoid treated groups and the 5 Gy native BM treated groups. (Figure 4B and Figure 4E,F). Consistent with hematological analysis, the osteo-organoid-derived cells exhibited enhanced regenerative capacity in the T cell subset chimerism in comparison to the native BM-derived cells (Figure 4C,D). The CD3+ T cell subset from the donor source achieved stable chimerism at 8 weeks post transplantation, while the B220+ B cell and CD11b+ myeloid cell subsets from the donor source achieved stable chimerism as early as 2 weeks after transplantation (Figure 4B–F). These data substantiated the role of osteo-organoid-derived cells from the donor in accelerating the recovery of T cell subsets in peripheral blood compared to the native BM-derived cells from the donor.
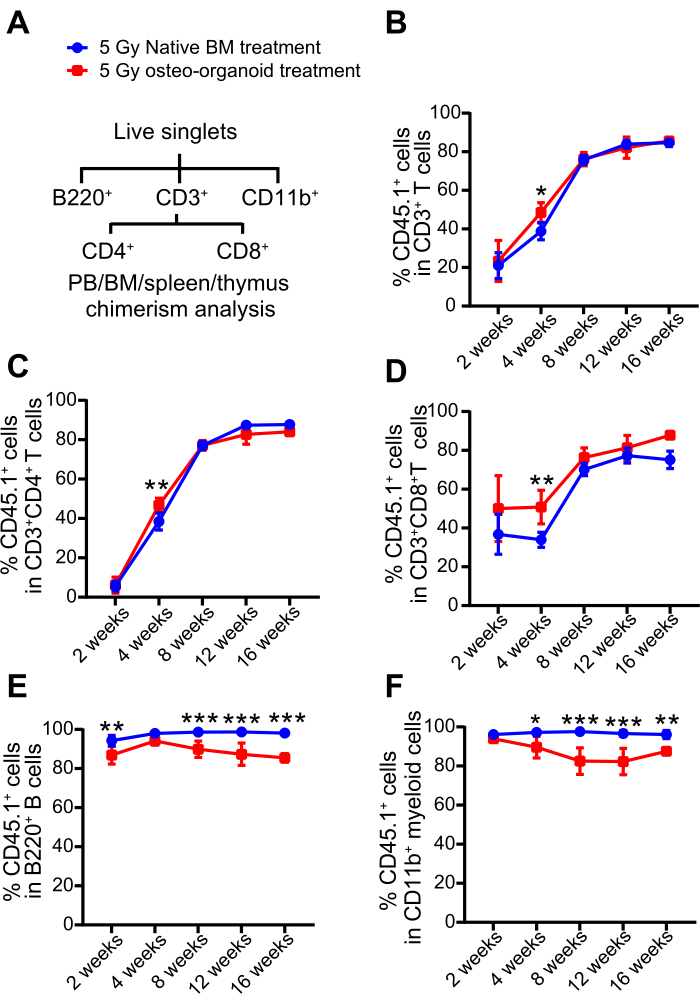
Figure 4: Reconstitution of the peripheral immune system in sublethally irradiated mice by osteo-organoid-derived cells. (A) The flow cytometric gating strategy for the chimerism analysis of recipient mice peripheral blood, BM, spleen, and thymus. (B–F) peripheral blood chimerism from the 5 Gy native BM-treated and 5 Gy osteo-organoid-treated groups was analyzed by the proportion of T cells, B cells, and myeloid cells at the indicated time points. Figure 4A–F was reproduced from Dai et al.10. n = 4 to 5 biological replicates. Data are presented as means ± SD. (B–F) Significant differences among groups were identified by two-way ANOVA followed by Bonferroni's post hoc test. *P < 0.05, **P < 0.01, and ***P < 0.001. Abbreviations: PB = peripheral blood; BM = bone marrow. Please click here to view a larger version of this figure.
To assess the regenerative capacity of transplanted cells in the irradiated mice's solid immune organs, we utilized the same flow cytometry gating strategy to analyze the chimerism of both native BM-derived and osteo-organoid-derived cells in the recipients' BM, spleen, and thymus (Figure 4A). At 16 weeks after cell therapy, the chimerism of T cells derived from donor osteo-organoid cells in recipients' BM was found to be increased, while the chimerism of B cells and myeloid cells was observed to be reduced, as compared to the chimerism of native BM cells from the donor (Figure 5A). Donor cells from both the 5 Gy osteo-organoid-treated group and the 5 Gy native BM-treated group exhibited pronounced chimerism in the spleen and thymus of the recipients, characterized by high levels of T cells, B cells, and myeloid cells (typically exceeding 80%) (Figure 5B,C). These findings provided evidence that osteo-organoid-derived and native BM-derived cells efficiently reconstituted the damaged immune system in mice exposed to irradiation.
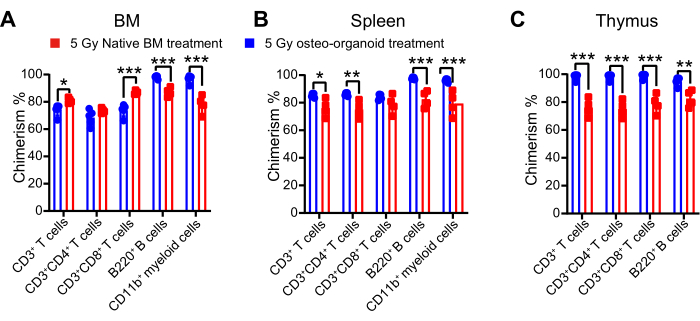
Figure 5: Reconstitution of the solid immune organ in sublethally irradiated mice by osteo-organoid-derived cells. (A–C) BM, spleen, and thymus chimerism from the 5 Gy native BM-treated and 5 Gy osteo-organoid-treated groups were analyzed by the chimerism rates of T cells, B cells, and myeloid cells at 16 weeks after transplantation. Figure 5A–C were reproduced from Dai et al.10. n = 4 to 5 biological replicates. Data are presented as means ± SD. Statistical differences among groups are identified by two-way ANOVA followed by Bonferroni's post hoc test. *P < 0.05, **P < 0.01, and ***P < 0.001. Abbreviation: BM = bone marrow. Please click here to view a larger version of this figure.
Supplemental Figure S1: Gating strategy used to identify HSPCs in the osteo-organoid. Single-cell suspensions from the osteo-organoid were subjected to flow cytometry for quantitative analysis of HSPCs. This figure was reproduced from Dai et al.10. Please click here to download this File.
Supplemental Figure S2: Gating strategy used to identify donor derived CD45.1+ cells in PB, BM, spleen, and thymus of CD45.2 recipients. Single-cell suspensions from PB, BM, spleen, and thymus of the CD45.2 recipients were analyzed via flow cytometry for the quantitative analysis of donor chimera. This figure was reproduced from Dai et al.10. Abbreviations: PB = peripheral blood; BM = bone marrow. Please click here to download this File.
