1. Program the Amaxa 96 well shuttle Nucleofector
- Open a new parameter file.
- Select the number of wells you will be using for standard transfection by dragging the cursor over the 96 well plate diagram. Use a minimum of 3 wells to pool for each experimental sample.
- Input the program code: in part1 select ‘FF’ and in part2 select ‘168’ from the pull down menus
- From Solution box select ‘Monocyte, human’
- Under Control Option select ‘standard’.
- Click on Apply.
- To include a no-transfection control, select further wells from the diagram as required and then choose ‘No Program Control’ from Control Option and click on Apply.
- Select any remaining unused wells on the plate diagram and click on Undefine.
2. Prepare DCs for transfection
- All work should be done under sterile conditions in a cell culture hood where possible. Prepare the nucleofection solution by adding 96-well supplement to Human Monocyte 96-well Nucleofector Solution in the ratio of 450 to 2025. Mix and allow to warm to room temperature. You will need 20μl of nucleofection solution per well. Make an excess of 10% to allow for pipetting error.
- Place the number of nucleocuvette modules you need into the nucleocuvette plate in the correct orientation ie inserting the first one in to rows 1 and 2.
- Determine the number of DCs needed for your experiment calculating on 500,000 cells per well and pellet by centrifuging at 400g for 10min. Carefully remove the supernatant.
- Resuspend the cells in the nucleofection solution by gently pipetting up and down a few times.
- Divide the correct volume of resuspended cells into eppendorfs labeled for the particular treatment eg GLO and RIG-I.
- Add 0.25μg siRNA per 500,000 cells and mix by pipetting. Use non-targeting siRNA in your no-transfection control sample.
- Pipette 20μl of the above mixtures into the nucleocuvette modules, according to your experimental layout, ensuring the liquid is delivered to the bottom of the well.
- Cover the nucleocuvette plate with the lid and tap the plate on a hard surface a couple of times to help ensure removal of air bubbles.
3. Transfect DCs
- Insert plate into the Nucleofector 96-well shuttle tray, click on Upload and then start.
- Follow progress of transfection process on the lap top display; a black cross on a green background signifies a successful transfection in that well, whereas a black bar on a red background means it was unsuccessful.
- On completion of the transfection process, remove the plate and add 80μl of DC growth medium to each well using a multichannel pipette.
- Incubate plate for 10 min at 37°C and 5% CO2.
- Transfer the 100μl volume from the nucleocuvettes into matrix tubes containing 100μl of pre-warmed DC growth medium, maintaining the correct orientation.
- Remove and discard those tubes where transfection did not occur.
- Incubate at 37°C and 5% CO2 for 24h or other desired time interval.
4. Infect cells with NDV
- Remove matrix tubes from the incubator to the cell culture hood and pool tubes for each experimental sample into eppendorfs
- Pellet the cells gently by spinning in a desk top centrifuge for 5 min and remove supernatant.
- Resuspend the cells in serum-free growth medium containing NDV at an MOI of 1 and incubate at 37°C and 5% CO2 for 45 min, with eppendorfs loosely covered in a sterile fashion.
- Add 900μl of DC growth medium, and re-incubate for 8-10 h.
5. Harvest cells
- Pellet cells by spinning eppendorfs in a desk top centrifuge and remove supernatant.
- Harvest cells for RNA or protein extraction according to your protocol.
6. Representative results:
Using our optimized protocol we transfected DCs with RIGI-targeting siRNA for 24 h and then infected the cells with NDV (a paramyxovirus detected by RIG-I) to stimulate the interferon response pathway. By qRT-PCR analysis we demonstrated knock down of the gene by 75% at the transcription level. We also observed a similar reduction in the expression of IFNβ, which is a downstream effector of RIG-I in the IFN signaling cascade. Furthermore, we observed that the expression of IFNβ in non-infected, control-transfected cells was not detectable while that of MxA, an IFNβ downstream response gene, was minimal (Figure 1A).
A second transfection of DCs with RIGI-targeting siRNA was performed using enough cells to include Western blot analysis. Where the RT-PCR results were similar to what we saw previously (62% and 66% knock down of RIG-I and IFNβ expression respectively) (Figure 1B), the Western blot probed for RIG-I revealed that the expression of this gene had been completely blocked (Figure 1C).
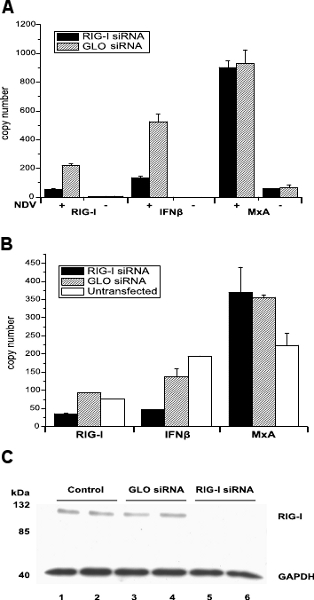
Figure 1. (A) MoDCs were transfected with either siRNA targeting RIG-I or nonspecific GLO siRNA and the effect on the expression of RIG-I, IFNβ and MxA determined by qRT-PCR. The transfection protocol used a monocyte-specific buffer and nucleoporation program FF168 (Lonza Walkersville Inc.) Following incubation for 24 h, transfected cells were either infected with NDV (+) or left uninfected (-). After a further incubation for 10 h, cells were harvested and RNA extracted Transcript levels represent the results of two replicate experiments. Taken from Bowles et al 16.
(B) MoDCs obtained from a different buffy coat as used for Figure 1A were again transfected with either RIG-I-targeting siRNA or nonspecific GLO siRNA using the same transfection protocol as above. An additional control using untransfected MoDCs was incorporated into the experiment. Following a 24 h incubation all cells were infected with NDV and incubated for a further 10 h before being harvested for both RNA and protein extraction. Transcript levels of RIG-I, IFNβ and MxA as determined by qRT-PCR represent the results of two replicate experiments. Taken from Bowles et al 16.
(C) Lysates from cells described in Figure 1B above were analyzed by Western blot. Lanes 1 and 2, lysates from untransfected cells; lanes 3 and 4, lysates from GLO siRNA transfected cells; lanes 5 and 6, lysates from cells transfected with RIG-I-targeting siRNA. Samples were probed for RIG-I and also GAPDH (as a loading control) and were run in duplicate. Taken from Bowles et al 16.
