Figure 1 shows a representative experiment of agroinfiltration with 7 different effectors (E1-E7) in potato and N. benthamiana. Cell death appears in the infiltrated leaf panels about 3 days after infiltration. The extent of the cell death phenotype needs to be compared with the controls. A mixture of the Agrobacterium strain AGL1(pVirG)23 containing pBINplus-R3a and pK7WG2-AVR3a was used as positive control24,25, while AGL1(pVirG) was used as negative control. Figures 1A and 1B show good examples of agroinfiltration in the potato genotype MaR8 (Mastenbroek R8)26 and N. benthamiana, respectively, which are very amenable for agroinfiltration. There is confluent cell death in the leaf panel coinfiltrated with positive control, while no cell death response occurs with negative control or pBINplus-R3a. In MaR8, two effectors AVR3a (E2) and AVR4 (E3) induce a cell death, while the other effectors (E1 and E4-E7) do not. In N. benthamiana, none of the tested effectors induce a cell death response. Figures 1C and 1D show examples of agroinfiltration in the wild potato Solanum berthaultii 483-1 and Solanum rechei 210-5, which are not well amenable for the agroinfiltration technique. In S. berthaultii 483-1, the leaf tissue shows a nonspecific necrosis to negative controls as well as to all tested effectors. In S. rechei 210-5, infiltrated leaf panels show very weak cell death response to positive control.
Figure 2 shows a range of scoring scales that can be used to quantify the response to agroinfiltrated Agrobacterium. Percentages of cell death are depicted on a scale 0-100%. Observed phenotypes range from macroscopically not visible symptoms (0%), through a range of intermediate responses displaying chlorosis and increasing levels of cell death, up to confluent cell death (100%).
Figure 3 shows a representative experiment after PVX agroinfection in potato. Normally, expanding cell death can be found at the sites about two weeks after tooth-pick inoculation. As shown in both figure panels, expanding cell death is present at the sites that were tooth-pick inoculated with pGR106-CRN2 (positive control using the effector crinkler Crn2), which is a general cell death inducing gene from P. infestans27. Apart from minor response to wounding, no expanding cell death is noted at the sites that were tooth-pick inoculated with the pGR106-empty (negative control). In Figure 3A, representing the potato genotype MaR3 (Mastenbroek R3), two pGR106-effectors (E1 and E2) did not induce cell death. In Figure 3B, a positive result of effector screening in the wild Solanum huancabambense 354-1 is presented; cell death response was observed to two pGR106-effectors (E1-E3, representing elicitins)28.
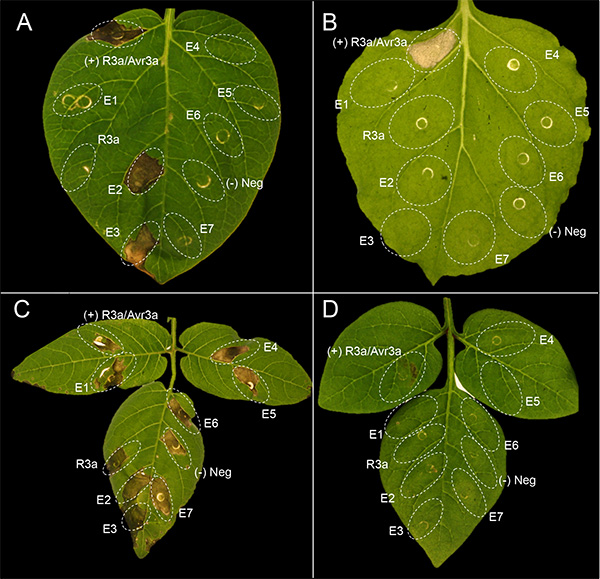
Figure 1. Examples of agroinfiltration in potato and N. benthamiana. Plants including (A) a potato genotype MaR8 (Mastenbroek R8), (B) N. benthamiana, (C) Solanum berthaultii 483-1 and (D) Solanum rechei 210-5 were infiltrated with a mixture of the Agrobacterium strain AGL1(pVirG) 23 containing pBINplus-R3a and pK7WG2-AVR3a (positive control), AGL1(pvirG) (negative control), pBINplus-R3a, and seven effectors (E1-E7). Click here to view larger image.
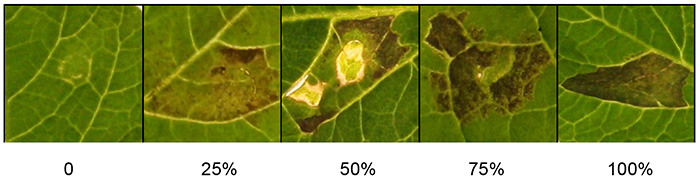
Figure 2. Quantification of agroinfiltration responses. The photograph shows representative scoring scales for cell death, ranging from 0% (no symptoms) to 100% (confluent cell death). Intermediate responses range from weak responses such as chlorosis to increasing levels of cell death. Click here to view larger image.
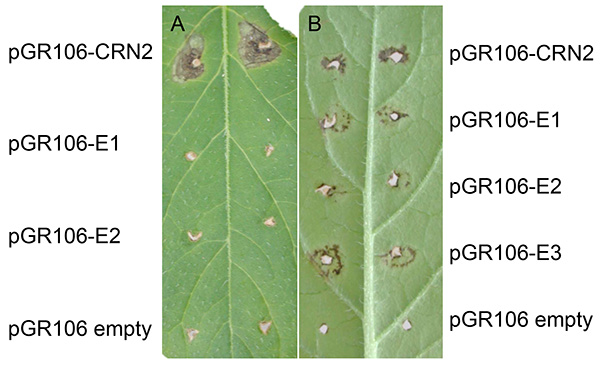
Figure 3. Examples of PVX agroinfection in potato. Potato genotype MaR3 (Mastenbroek R3) (A) and Solanum huancabambense 354-1 (B) tooth-pick inoculated with pGR106-CRN2 (positive control), pGR106-empty (negative control) and pGR106-E1-2 (effectors), or pGR106-E1-E3, respectively. Click here to view larger image.
Table 1. YEB medium
| 1 L | distilled H2O |
| 5 g | sucrose |
| 5 g | beef extract |
| 5 g | bacteriological peptone |
| 1 g | yeast extract |
| 2 ml | MgSO4 (1 M) |
Table 2. Vectors and strains used for agroinfiltration and PVX agroinfection. Several binary vectors can be used. Vectors in the list below allow high expression of the candidate genes and have worked well in our hands. We prefer using the Agrobacterium strain GV3101(pMP90)29 di N. benthamiana and find AGL130 containing the helper plasmid pVirG (pBBR1MCS-5.virGN54D)23 more suitable in potato31. Additional strains have been analyzed in other groups on various model plants including N. benthamiana but not in potato32.
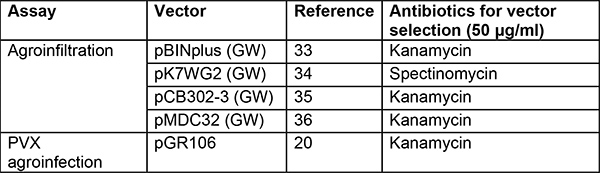
GW: gateway version36
Table 3. MMA medium.

Adjust the pH to 5.6.
