Major aim of this study is to demonstrate a method for easy and accurate identification of gene silenced N. benthamiana plants that are compromised for nonhost resistance. There are four major steps in this methodology. First step is to individually silence large number of genes using TRV-VIGS. We had silenced about 5,000 genes6,18 over a period of about 1.5 years using the protocol depicted in Figure 1. Some of the gene silenced plants showed various phenotypic alterations including stunted growth, yellowing and photo-bleaching phenotypes (Figure 2).
Second step is the identification of gene silenced plants showing susceptibility to nonhost pathogen(s). We used GFPuv expressing bacteria and tracked their growth in planta (Figures 3A-3C). In order to demonstrate the process of identification of the gene silenced plants susceptible to nonhost pathogen(s), we used three nonhost pathogens, P. syringae pv. tomato T1, P. syringae pv. glycinea and X. campestris pv. vesicatoria. As expected, non-silenced (vector control) N. benthamiana plants inoculated with these pathogens did not grow as they could not overcome the plant immunity. However, P. syringae pv. tomato T1 formed green fluorescent colonies on NbSGT1 gene (a gene implicated in nonhost resistance25) silenced plants (Figure 3D). These data demonstrate the process of identifying gene silenced plants compromised for nonhost resistance. This protocol can be directly applied to forward genetics screening. For example, the representative results taken from one of our previous forward genetics screen (Figure 4) shows two independent gene silenced plants with varying degree of P. syringae pv. tomato T1 growth (Figure 5).
Third step is the confirmation of bacterial growth in the gene silenced plants identified from the screen by quantifying the bacterial growth. An example for estimation of in planta bacterial growth in the NbSGT1 gene silenced plants is shown in Figure 3E. Results showed that in planta growth of P. syringae pv. tomato T1 was much higher in the NbSGT1 silenced plants compared to non-silenced (TRV::00) control plants (Figure 3E). Loss of nonhost resistance due to a particular gene silencing could be either partial or complete. Depending on this, the extent of nonhost pathogen growth on the gene silenced plants can vary.
Fourth step is sequencing the insert in TRV2 vector. BLAST is performed using this sequence to identify the gene and its details.
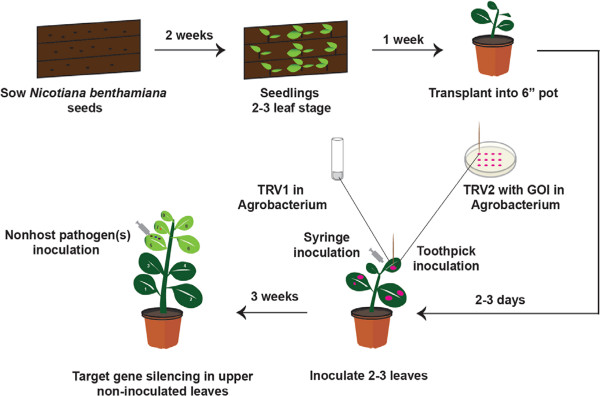
Figure 1. Cartoon showing the steps involved in silencing a large number of genes using TRV-based VIGS. Three week old N. benthamiana plants are transplanted into individual pots and allowed to grow for a few days. Agrobacterium carrying TRV1 construct is infiltrated into three to four leaves using needless syringe. Later, 96 cDNA clones grown on LB agar plate are individually taken using a toothpick and pricked on to the TRV1 inoculated spot of respective plants. About 3 weeks after inoculation, target gene silencing occurs in the top non-inoculated newly grown leaves. These leaves (5th to 8th) are used for testing the growth of nonhost pathogen(s).

Figure 2. VIGS-mediated forward genetics screening in N. benthamiana shows various changes in plant phenotype. Three-week old N. benthamiana plants independently inoculated with respective TRV constructs as descried in Figure 1 as a part of forward genetics screen are shown. Photographs were taken at 21 days after TRV inoculation.
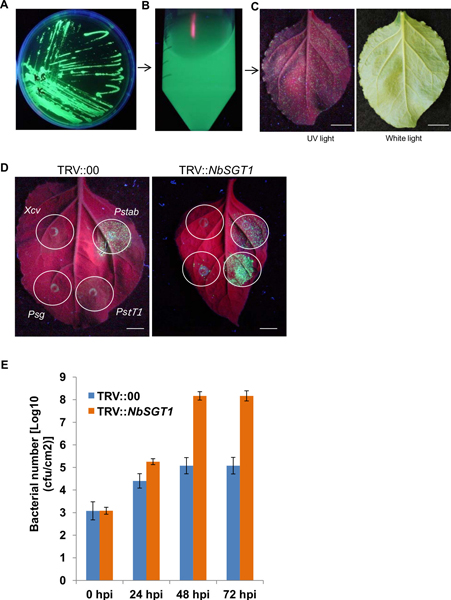
Figure 3. Green fluorescence from GFPuv expressing bacterial pathogens and its use for identification of genes involved in nonhost resistance. Pseudomonas syringae pv. tabaci, a host pathogen, carrying GFPuv construct streaked on KB-medium plate shows green fluorescence under long wave UV light in the dark (A). These bacteria are grown on KB medium and re-suspended in water (B) and inoculated on to N. benthamiana leaves (abaxial side, 1 x 104 cfu/ml). At 3 days post inoculation (dpi) bacterial colonies are viewed as green fluorescent spots by naked eye from abaxial side of leaf epidermal cells (C, left). The same leaf pictured under normal light (visible range) is also shown (C, right). GFPuv expressing P. syringae pv. tabaci (Pstab, 1 x 104 cfu/ml) and three nonhost pathogens, P. syringae pv. tomato T1 (PstT1, 1 x 104 cfu/ml), P. syringae pv. glycinea (Psg, 3 x 105 cfu/ml) and Xanthomonas campestris pv. vesicatoria (Xcv, 3 x 104 cfu/ml) are inoculated on to the abaxial side of leaves from vector control (TRV::00; D, left) and NbSGT1 gene silenced plants (D, right). Bacterial colonies of Pstab are seen on both leaves and those of PstT1 are seen only on the NbSGT1 gene silenced leaves. PstT1 growth in NbSGT1 gene silenced leaves quantified at 24, 24 and 72 hr post inoculation (hpi, E) is shown in the graph. Click here to view larger figure.
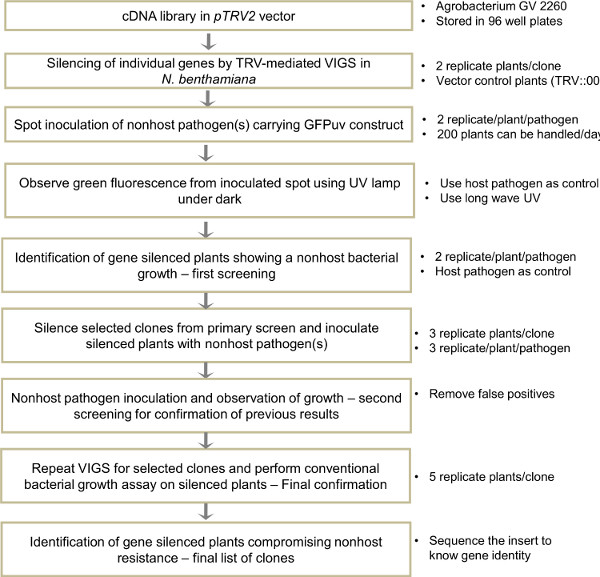
Figure 4. Overview of protocol followed for VIGS-mediated forward genetics screening for identification of genes involved in nonhost resistance. A cDNA library in TRV2 vector constructed using RNA from nonhost pathogen(s) inoculated N. benthamiana leaves can be used for screening. Individual clones from the cDNA library are silenced in N. benthamiana by TRV-VIGS. Respective GFPuv expressing nonhost bacterial pathogen(s) is inoculated on to the gene silenced plant leaves. Between 2 and 4 dpi the inoculated spots are observed using a UV lamp in the dark and the plants showing nonhost pathogen growth are identified. After this preliminary screening, the selected clones are silenced again and the nonhost pathogen growth is reconfirmed. This second level screen is done to remove the false positives. Later, confirmed clones are silenced for a third time and respective bacterial growth is estimated. Inserts of respective clones in the TRV2 vector are sequenced and gene information is identified. Simultaneously HR assay can also be performed on the gene silenced plants to identify genes contributing to HR induced against type-II nonhost pathogens.
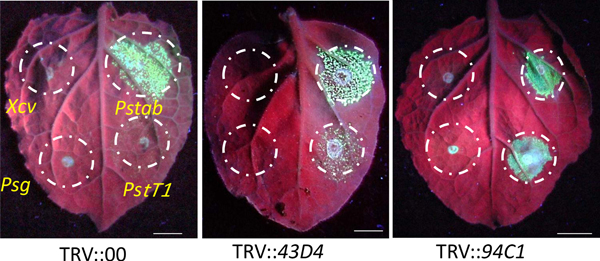
Figure 5. Nonhost bacterial pathogen growth in gene silenced plants that were identified as part of a forward genetics screen. Pictures from a forward genetics screening that identified genes involved in nonhost resistance of N. benthamiana plants against PstT1, Psg and Xcv. Photographs taken for two representative gene silenced plants at 3 dpi after inoculation are shown here. Bacterial concentrations used for inoculation are described in Figure 3. 43D4 or 94C1 indicates the 96-well plate number followed by the well number.
