The Casp3 assay gives clear evidence of apoptotic cells from the 4-cell stage to 32 hpf. Figure 2A shows that injection of embryos with high concentrations of mRNA encoding potent pro-apoptotic bh3-only proteins, such as Bim or Noxa, quickly gives rise to active-Caspase 3 by the 4-cell stage of development. Embryonic death subsequently occurs within 1 hr4. Note that intensity of fluorescence is moderate, but results are conclusive. At older stages, fluorescence intensity is more robust. For instance, Figure 2B shows bright active-Caspase 3-positive cells in sphere-stage embryos that have been injected with moderately toxic levels4 of mRNA encoding the pro-apoptotic bh3-only gene puma. Messenger RNA encoding the anti-apoptotic bcl-2 gene was coinjected to prove that injection of puma mRNA induces apoptosis through the intrinsic, mitochondria-mediated pathway.
IR-induced apoptosis in zebrafish requires p53-mediated transcription of bh3-only genes such as puma5-6. The delay in IR-induced apoptosis in Figure 3 is consistent with the requirement for p53-mediated transcription.
The Casp3 protocol should give rise to a robust signal-to-noise ratio. Figure 4 compares the optimal Casp3 assay result to two common suboptimal results that arise when critical steps are compromised (see Discussion): A weak signal-to-noise ratio due to background fluorescence in animal tissue and yolk fluorescence that distracts from the signal-of-interest.
Colabeling experiments using transgenic GFP-expressing lines approximate tissues undergoing apoptosis (Figure 5).
Figure 1. Simplified schematic of the Casp3 protocol.
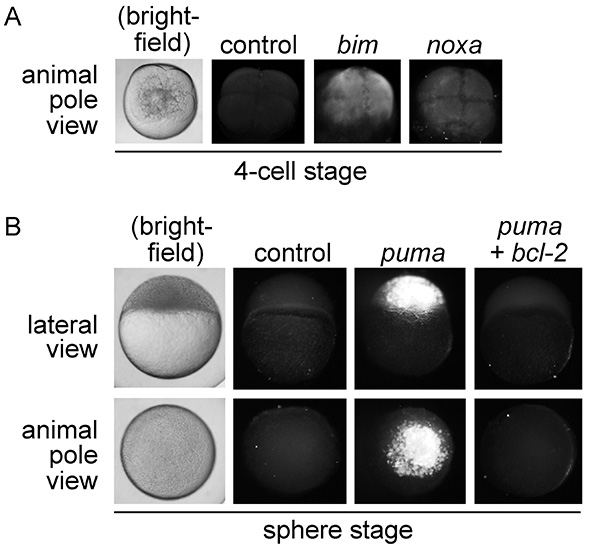
Figure 2. (A) Analysis of apoptosis in 4-cell stage embryos. Embryos were injected at the one-cell stage of development with 50 pg of mRNA encoding egfp (control) or the pro-apoptotic bh3-only genes bim and noxa. At the 4-cell stage (approximately one hr-post-injection, hpi), embryos were analyzed by immunofluorescence to detect activated Caspase 3. Bim and noxa mRNA induce rapid apoptosis in zebrafish embryos immediately following injection which, if embryos are left to develop, leads to death by 2 hpi4. A brightfield image of a wild-type 4-cell stage embryo is shown for comparison. (B) Analysis of apoptosis in sphere-stage zebrafish embryos. Embryos were injected at the one-cell stage of development with mRNA encoding either 50 pg egfp (control), 5 pg puma plus 45 pg egfp (puma), or 5 pg puma plus 45 pg bcl-2 (bcl-2). At sphere stage (approximately 4 hpi), embryos were analyzed by the Casp3 assay. Both lateral and animal pole views are shown to visualize apoptotic cells. Rescue of puma-induced apoptosis by bcl-2 indicates that injection of puma mRNA induces apoptosis specifically through the intrinsic, mitochondria-mediated pathway. A brightfield image of a wild-type sphere-stage embryo is shown for comparison.
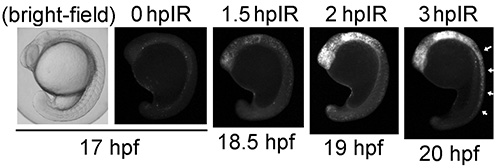
Figure 3. Time course of ionizing radiation (IR)-induced apoptosis between 17-20 hpf. Zebrafish embryos were irradiated with 8 Gy IR at 17 hpf and analyzed at 0, 1.5, 2, and 3 hr-post-IR (hpIR) by the Casp3 assay. Arrows at 20 hpf show accumulation of apoptotic cells in the spinal cord at 3 hpIR. A brightfield image of a 17-hpf embryo is shown for comparison.
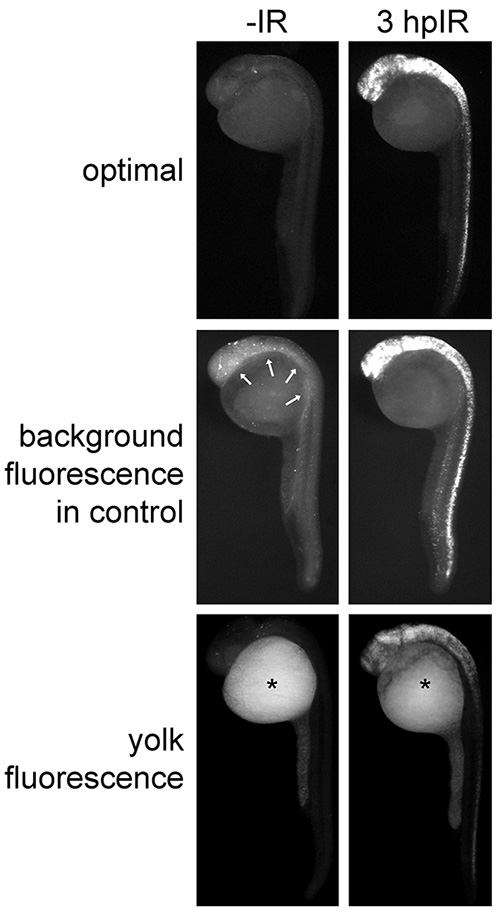
Figure 4. Optimal and sub-optimal Casp3 immunofluorescence results on 27-hpf embryos. Zebrafish embryos were irradiated (or not) at 24 hpf with 15 Gy and analyzed by the Casp3 assay. The “optimal” group shows ideal results in which apoptotic cells brightly fluoresce, and nonapoptotic cells and the yolk show minimal background fluorescence. Arrows in the “background fluorescence in control” group denote excessive background fluorescence in the animal tissue of control embryos. Asterisks in the “yolk fluorescence” group demonstrate excessive nonspecific fluorescence in the yolk.
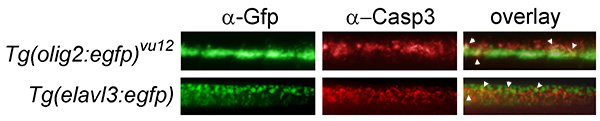
Figure 5. Colabeling experiments approximate tissue specificity of IR-induced apoptosis. The Tg(olig2:egfp)vu12 transgenic line labels motor neurons and oligodendrocytes17, and the Tg(elavl3:egfp) line labels all differentiating neurons18. Each transgenic line was irradiated with 15 Gy at 24 hpf and analyzed 3 hr later using both mouse anti-GFP and rabbit anti-active-Caspase 3 primary antibodies, followed by green- and red-fluorescing secondary antibodies, respectively. A magnification of the spinal cord (located dorsal to the end of the yolk extension of 27-hpf, 3-hpIR embryos) is shown. Apoptotic cells appear to be partially localized to both Tg(elavl3:egfp) and Tg(olig2:egfp)vu12 populations (see arrowheads in overlay), suggesting that neurons and oligodendrocytes are sensitive to IR-induced apoptosis.
