The steps diagrammed in Figure 1 indicate the appearance of the embryo while attached to the vitelline membrane (A) and demonstrate the proper method to separate the embryo from the yolk (B). The embryo can be identified by the zone of junction, which is much lighter than the vitelline membrane. The embryo itself is often difficult to distinguish until the yolk is cut away. Once the embryo is dissected from the egg, it can be fixed or flash frozen for future use. If an in situ hybridization is planned for the dissected embryo, it is necessary to remove the vitelline membrane that is adhered to the embryo via the zone of junction. Figure 2 illustrates the improved visibility of the embryo once this membrane is removed (C), and the proper way to peel the vitelline membrane (A, B). After dissection and fixation, whole mount in situ hybridization was performed as seen in Figure 3 (A, A’, B, B’) and Figure 4 (A, A', B, B’) and Figure 5 (A, B, C) to detect differences in orthodenticlehomeobox 2 (Otx2) expression in embryos developmentally exposed to low doses of methylmercury. Figure 5 shows sense probe results, demonstrating lack of background. In Figure 4, despite being dissected from the egg at the same time point, the embryo developmentally exposed to methylmercury (MeHg) progressed to stage 522 (B, B’), while the control embryo developed to stage 622 (A, A’). The group of embryos dissected and shown in Figure 4 were collected from the nest and taken from the incubator at the same times. Although some natural variation is present in development, based on previous dissection data, it is unlikely that temperature fluctuations in the incubator would cause only the 2.4 ppm methylmercury embryos to be developmentally delayed. The differences in stages indicate changes in cell proliferation in embryos developmentally exposed to methylmercury.
Before dissection, EdU was injected into a day 2 egg and allowed to incubate overnight. After dissection and fixation of the stage 1622 embryo, EdU was visualized using “click” chemistry, allowing detection of proliferating cells as seen in Figure 6 (A, B, C). It is important to carefully monitor time points when placing eggs in the incubator and during dissections, as exposure to methylmercury or performing the EdU assay may disrupt developmental progression. The earliest injection was performed on day 0, which was the day of collection as specified in step 1.3. This embryo was dissected approximately 38 hours later (stage 722). The survival rate was found to be approximately 90% (same rate as control embryos) as long as the injection amount was under 478 nl.
This dissection methodology also allows for high quality RNA extraction. After dissecting stage 1622 embryos, an RNA extraction was performed according to manufacturer’s protocol with no optimization required, as seen in Figure 7. The removal of the vitelline membrane was unnecessary for RNA extraction and later qRT-PCR applications.
Note: All embryo figures are oriented so that the anterior and posterior regions are at the top and bottom of the images, respectively.
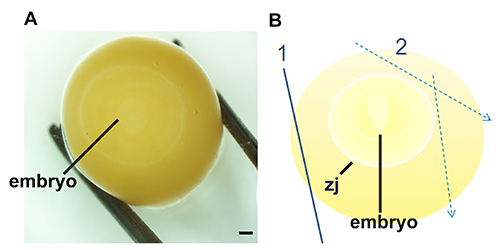
Figure 1. Procedure for locating and dissecting zebra finch embryos, stages 1-1022. Locate embryo by gently rolling the yolk until the faint white disk is apparent (A). Once the embryo is located at the center of the yolk, the yolk is dissected in a stepwise fashion (B) where the first cut relieves pressure of the vitelline membrane (1) and subsequent cuts (2) border the zone of junction (zj) which is adhered to the vitelline membrane. Scale bars represent 1 mm.
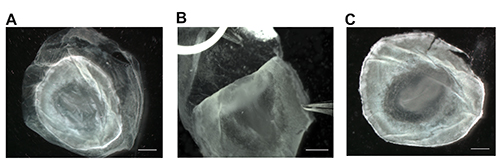
Figure 2. Removal of vitelline membrane and visibility of embryonic structures. Following removal of the embryo from the yolk, place embryo in a Petri dish containing 4% PFA. If an in situ hybridization needs to be performed, visibility of the embryonic structures is essential and can be achieved by removing the vitelline membrane (A). Grip the vitelline membrane with extra fine tipped forceps and gently peel it away from the embryo by handling the embryo directly at the outermost edge, if necessary (B). Vitelline membrane removal increases clarity of embryonic structures, and allows embryos to be imaged or processed with in situ hybridization (C). Scale bars represent 1 mm. Please click here to view a larger version of this figure.
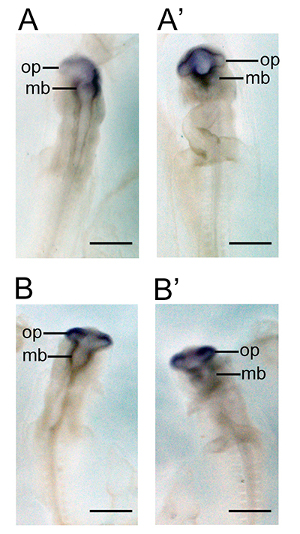
Figure 3. Whole mount in situ hybridization performed on zebra finch embryos developmentally exposed to methylmercury. Expression patterns of orthodenticlehomeobox 2 (Otx2) were characterized in embryos exposed to 0.0 ppm methylmercury (A, A’) and 2.4 ppm methylmercury (B, B’) via parental diet. The dorsal (A) and ventral (A’) expression of Otx2 is visible throughout the midbrain and optic vesicles during stage 1222. The treatment group embryos were dissected at the same time point, but were developmentally delayed as seen in the dorsal (B) and ventral (B’) view of head structures, which are characteristic of stage 1122. Abbreviations: mb, midbrain; op, optic vesicle. Scale bars represent 1 mm.
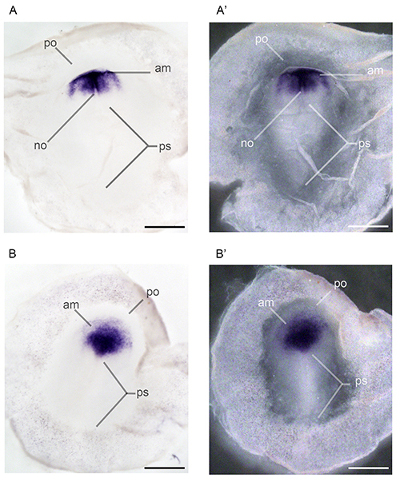
Figure 4. Whole mount in situ hybridization performed on zebra finch embryos developmentally exposed to methylmercury. Expression patterns of orthodenticlehomeobox 2 (Otx2) were characterized in stage 622 embryos exposed to 0.0 ppm methylmercury (A, A’) and stage 522 embryos exposed to 2.4 ppm methylmercury (B, B’) via parental diet. Abbreviations: am, anterior margin of mesoderm; no, notochord, notochord mesoderm; po, proamnion, anterior blastopore; ps, primitive streak22. Scale bars represent 1 mm.
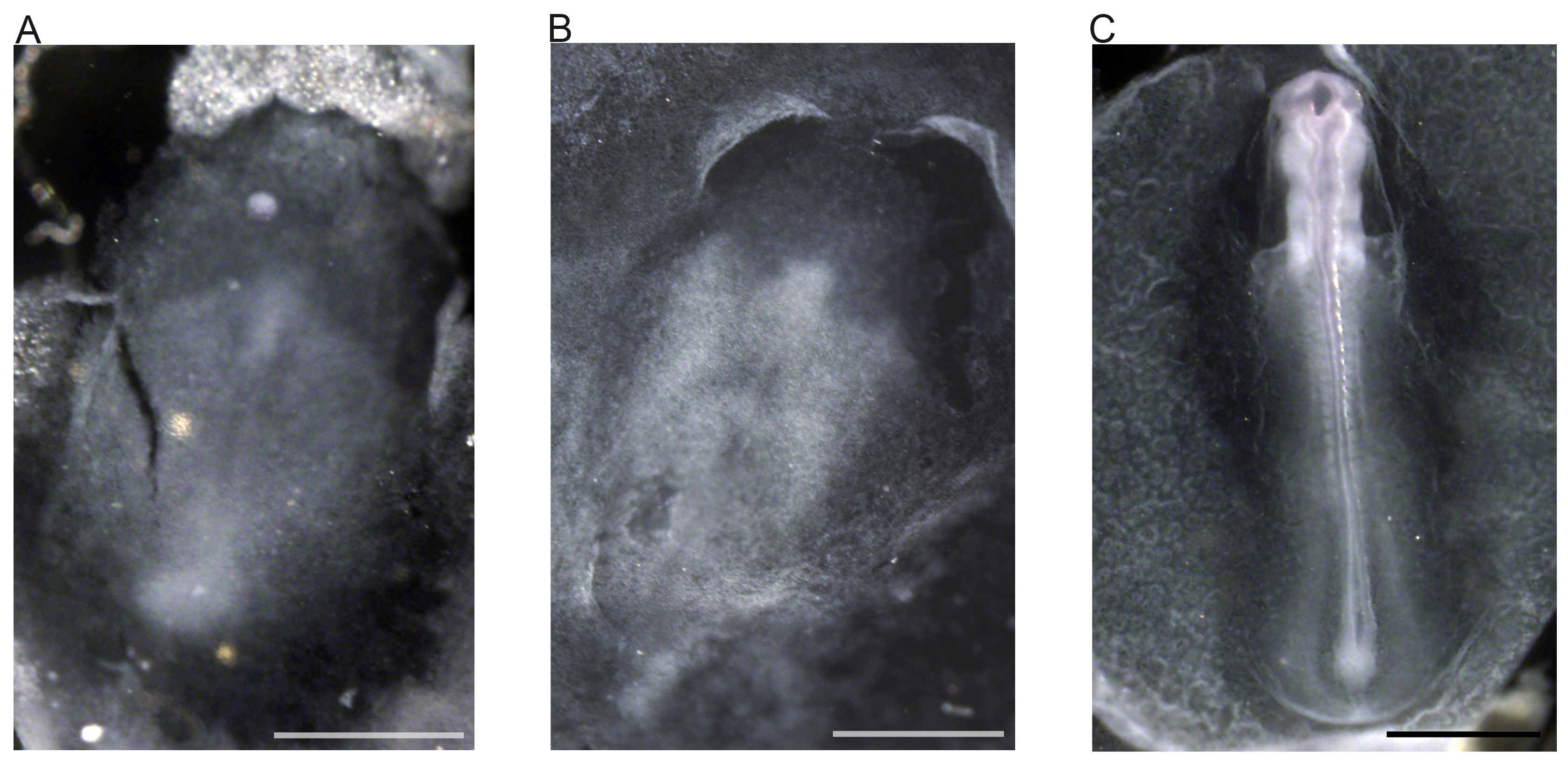
Figure 5. Whole mount in situ hybridization performed on zebra finch embryos using sense probe. (A) Stage 522 embryo. (B) Early stage 622 embryo. (C) Stage 1122 embryo. Scale bars represent 1 mm.
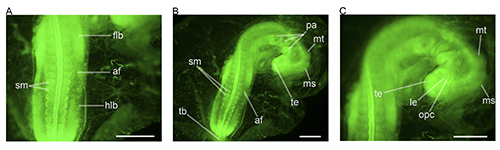
Figure 6.EdU incorporation and detection in zebra finch embryos. EdU “click” chemistry was used to detect proliferating cells in a stage 1622 embryo (A, B, C). EdU is incorporated into the DNA in the place of thymidine26,27 and is detected using click chemistry27. Proliferation is clearly visible in the lateral edges of the somites, and the tailbud. Panel A shows proliferation occurring exclusively in the posterior of the embryo and also shows individual proliferative cells. Panel B shows the proliferative locations in the whole embryo. Panel C shows the anterior region, and shows the highly proliferative telencephalon (te) in greater detail. Abbreviations: af, amniotic fold; flb, forelimb bud; hlb, hindlimb bud; le, lens vesicle; ms, mesencephalon; mt, metencephalon; opc, optic cup; pa, pharyngeal arch; sm, somite mesoderm; tb, tailbud; te, telencephalon. Scale bars represent 1 mm. Please click here to view a larger version of this figure.
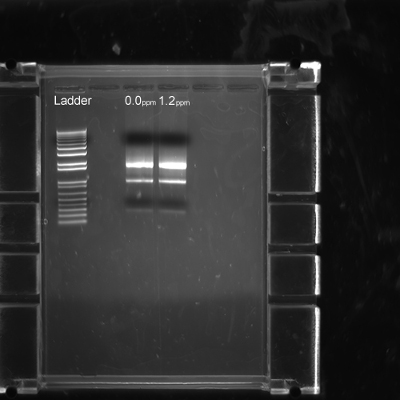
Figure 7. Quality of RNA extracted from dissected zebra finch embryos. Control (0.0 ppm) and 1.2 ppm methylmercury embryos were dissected and flash frozen as described in Step 3.8. Each lane shows RNA extracted from two homogenized embryos from each treatment group.
