The IFN-γ ELISpot protocol detailed above was developed and optimized in our laboratory to measure the magnitude and the quality of cell mediated immune responses directed against VZV4. Diverse sources of VZV antigen can be used for the stimulation step. These include: a) commercially available detergent inactivated extracts from VZV infected Vero cells18; b) pools of overlapping synthetic peptides from specific VZV encoded proteins, including IE6315 and ORF419; c) live attenuated varicella zoster vaccine20; and d) UV inactivated preparations of VZV antigens from the supernatant of disrupted VZV infected MRC cells14. In our case, dilutions of live attenuated VZV vaccine were preferred over cell culture derived VZV in order to insure uniformity in antigen source and preparation and to limit culture and manipulation of live wildtype VZV strains in a clinical setting where nosocomial transmission is a serious concern10. In addition, γ irradiation was preferred over UV inactivation21 because of the comparative precision with which doses of γ radiation can be delivered and because, unlike UV, γ-rays efficiently penetrate sealed VZV containing vials. Irradiation of viral stocks led to consistently higher estimates of the frequency of VZV specific IFN-γ producing cells in PBMC samples obtained from a healthy donor at all dilutions of VZV antigen tested (Figure 1). This is possibly the result of the mitigation of cytopathic effects induced by live attenuated VZV in cell culture.
To determine which dilution VZV antigen yielded maximal readouts in terms of numbers of SFU per 106 PBMC, IFN-γ ELISpot was performed using two-fold dilutions of γ irradiated live-attenuated VZV vaccine and PBMC samples obtained from a group of immunocompetent children aged between 9 months and 12 years who had a documented history of varicella and/or a positive serology for VZV (n = 50). Study subjects were enrolled at the Infectious Diseases Clinic and Special Immunology Clinic of CHU Sainte-Justine between June 2007 and September 2009. Statistically significant differences were observed between median frequencies of IFN-γ producing cells that were obtained using the 1:200, 1:400 and 1:800 dilutions of VZV antigen (p < 0.0001, Kruskal-Wallis test) (Figure 2). These disparities were accounted for by the difference between the 1:200 dilution (median = 115.0 SFU per 106 PBMC; interquartile range [IQR] = 75.0 – 232.5 SFU per 106 PBMC) and the 1:800 dilution (median = 50.0 SFU per 106 PBMC; IQR = 20.0 – 105.0 SFU per 106 PBMC), and between the 1:400 dilution (median = 92.5 SFU per 106 PBMC; IQR = 52.5 – 177.5 SFU per 106 PBMC) and the 1:800 dilution (p < 0.001 and p < 0.01, respectively, Dunn’s multiple comparisons test) (Figure 2). For this reason, the 1:200 dilution of γ irradiated live-attenuated VZV vaccine was selected for use in routine measurements of VZV specific cell mediated immune responses.
To determine whether IFN-γ production resides in CD4+ and/or CD8+ T lymphocyte subsets, PBMC samples obtained from a healthy donor were subjected to depletion of CD4+ or CD8+ cells using anti-CD4 or anti-CD8 antibody coupled magnetic beads according to the manufacturer’s instruction. In concordance with previously published reports22,23, depletion of CD4+ cells resulted in a striking reduction in the frequencies of IFN-γ producing cells that were obtained using the 1:100, 1:200 and 1:400 dilutions of VZV antigen, while depletion of CD8+ cells did not lead to such a reduction and positive controls (stimulation with anti-CD3 monoclonal antibody) were not similarly affected (Figure 3). These results suggest that the majority of cells that produce IFN-γ in response to stimulation with VZV antigens are CD4+ T cells. Alternatively, these results could stem from depletion of CD4+ antigen presenting cells (i.e. monocytes, dendritic cells) from the PBMC pool, leading to reduced levels of presentation of VZV antigens to responder CD8+ T lymphocytes. The use of irradiated VZV is unlikely to have contributed to the skewing of the response towards CD4 dominance, as similar results were obtained by other groups using different techniques and antigen sources14,21,22. A low magnification micrograph of representative ELISpot wells is presented in Figure 4.
| Problem/issue | Possible causes | Remedy/solution |
| High background/Poorly-defined or confluent spots | High numbers of dead cells. | Assess cell viability before culture set-up and stimulation. |
| Inadequate membrane pre-wetting steps. | Make sure to respect pre-wetting time and that membrane turn to gray after 1 min. 35% ethanol should be prepared immediately before use. | |
| Plates moved during the incubation period. | Do not move plates to avoid poorly-defined snail trail spots. | |
| Wash steps. | Manual wash is more efficient than plate washers. | |
| Formation of protein aggregates. | Filter both capture and detection antibodies to reduce background and false positive spots. | |
| Secondary and detection antibody concentration. | Adjust biotinylated secondary/detection antibody concentration to reduce background. | |
| Cold developing reagents. | Bring substrate to room temperature to avoid underdevelopment. | |
| No spots/Blank wells | Poorly defined spots. | Do not stack plates during incubation period to have a homogeneous temperature in the different wells. |
| Stimulation substrate not well prepared. | Bring the peptide mixture aliquot to room temperature for 15 min to avoid crystal formation. | |
| Cell clumping. | Resuspend cells into single-cell suspension to avoid underestimation of spot-forming units. | |
| Cells not stimulated appropriately. | Use a polyclonal activator as a positive control. | |
| Inhibition of enzyme reaction by tween-20. | Wash plates with PBS before adding BCIP/NBT substrate solution. | |
| Wrong antibody pairs. | Make sure that the capture and detection antibodies react with different antigenic epitopes. |
Table 1. Additional troubleshooting.
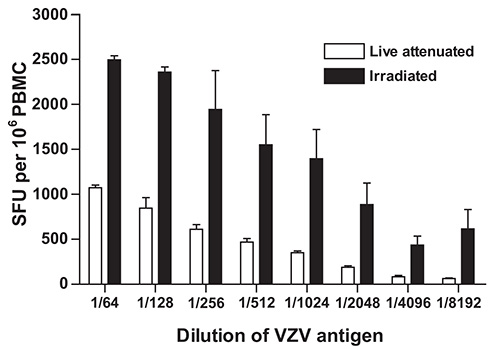
Figure 1. Effect of γ-irradiation of live inactivated VZV vaccine on the frequency of IFN-γ producing cells as determined using IFN-γ ELISpot. VZV specific IFN-γ ELISpot was performed as described under Protocol Text using PBMC from a control volunteer and serial dilutions of γ irradiated (10,000 Gy) live attenuated VZV vaccine as antigen. Data represent the mean of duplicate measurements and error bars represent the standard error of the mean. PBMC: peripheral blood mononuclear cells; SFU: spot forming units; VZV: varicella zoster virus.
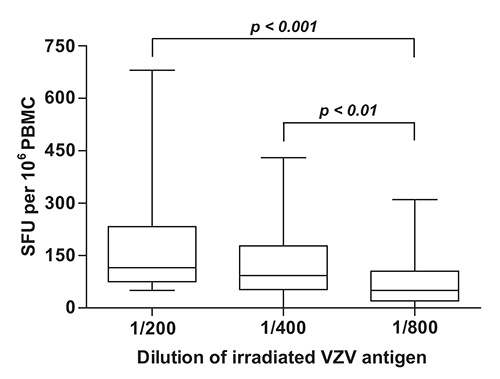
Figure 2. Quantification of VZV specific cell mediated immune responses in children with documented evidence of previous infection with VZV. VZV specific IFN-γ ELISpot was performed as described under Protocol Text using PBMC isolated from subjects with a history of varicella (n = 50) and serial dilutions of γ irradiated (10,000 Gy) live attenuated VZV vaccine as antigen. Horizontal lines represent the median, boxes represent interquartile range (IQR), and whiskers represent the range of values. PBMC: peripheral blood mononuclear cells; SFU: spot forming units; VZV: varicella zoster virus.
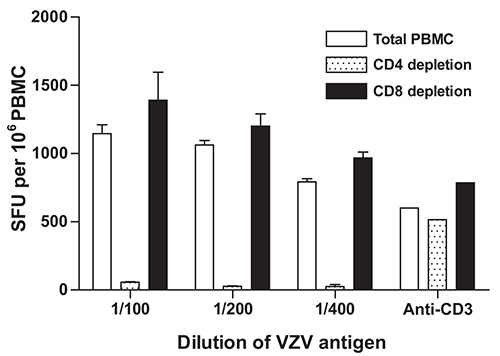
Figure 3. Effect of depletion of CD4+ or CD8+ cells on the frequency of IFN-γ producing cells as measured using IFN-γ ELISpot. Depletion of CD4+ or CD8+ cells and VZV specific IFN-γ ELISpot were performed as described under Protocol Text using PBMC from a control volunteer and serial dilutions of γ irradiated (10,000 Gy) live attenuated VZV vaccine as antigen. Data represent the mean of duplicate measurements and error bars represent the standard error of the mean. PBMC: peripheral blood mononuclear cells; SFU: spot forming units; VZV: varicella zoster virus.
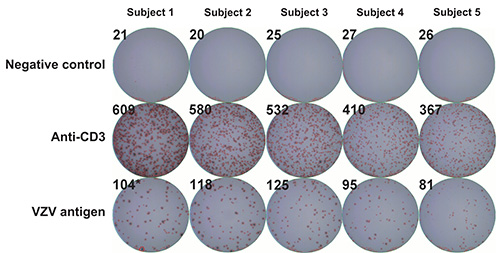
Figure 4. Representative ELISpot wells. VZV specific IFN-γ ELISpot was performed as described under Protocol Text using PBMC from five control volunteers and serial dilutions of γ irradiated (10,000 Gy) live attenuated VZV vaccine as antigen. First row: negative controls (AIM-V medium supplemented with 2% (v/v) IHS); second row: positive controls (anti-CD3 monoclonal antibody); third row: VZV antigen. Numbers in the upper corners represent SFU per well as measured using an automated spot counter. PBMC: peripheral blood mononuclear cells; SFU: spot forming units; VZV: varicella zoster virus; IHS: inactivated human serum.
