The present results show that the high throughput pipeline to study S. epidermidis and M. marinum infection has been successfully established and that may be extended to other infection models. Firstly, the use of the large breeding vessel (Figure 1A), based on the published system by Adatto et al. (2011)11, enables to generate large numbers of synchronous eggs in single events affording a high control of the spawning process. Next to be able to inject large number of embryos in a short period of time, an improved version of the previously developed automated micro-injection system7 was used (Figure 1A). To asses which is the best developmental stage for yolk infection, injections with S. epidermidis and M. marinum were performed at all the different stages between 1 and 512 cell stage, according to the description made by Kimmel et al. (1995)12.
Injections with 100 cfu S. epidermidis between the 16 and 128 cell stage provided the best infection pattern (Figure 2). The bacteria proliferated inside the yolk for 3 days and spread into the body from 3 dpi onwards. Performing injections before the 16 cell stage led to high mortality from 4 dpi, and injection after the 256 cell stage showed mainly bacterial growth inside the yolk with hardly any bacteria spreading inside the body of the embryo. Quantification of bacterial burden was performed by fluorescence intensity analysis using the large particle flow cytometer as described by Veneman et al. (2013)8 (Figure 3).
Observations showed that the optimal developmental stage for injection of 30 cfu M. marinum injection is between 16 to 128 cell stage for the E11 strain (Figure 4A) and between 16-64 cell stage with the more virulent M strain (Figure 4B). Embryos injected at these stages showed bacterial growth inside the yolk and spreading of the bacteria through the embryo (Figure 7). Infection with both strains at earlier stages presented nonspecific generalized bacterial growth leading the embryos to die after 4 dpi. On the other hand, in embryos injected at later stages bacterial burden was restricted to the yolk.
Next, presorting with large particle flow cytometer (Figure 1B) generated large homogenous groups of infected fish excluding non- or highly infected embryos (Figures 5A and 6A). After presorting, M. marinum infected embryos were treated with Rifampicin, a first line antituberculosis drug. Previous studies demonstrated that treatment with Rifampicin at a dose of 200 μM efficiently reduces M. marinum infection in zebrafish7,13. Taking advantage of the large number of homogenously infected embryos generated with the high throughput setup, treatment with different doses was performed. Embryos infected with M. marinum M strain and treated for 48 hr with 12, 24, and 200 μM Rifampicin showed to reduce efficiently mycobacterial infection in a dose dependent manner (Figure 5B). In view of the efficient reduction of the infection using Rifampicin at a dose of 200 μM this concentration was used for the future experiments. In line with the previous result, studying bacterial burden progression using M. marinum E11 strain a significant reduction 24 hr and onwards after treatment with 200 μM Rifampicin was observed (Figure 6B).
Furthermore, if high magnification imaging is required of these embryos, they can be displayed automatically in 96-well plates (Figure 1C), from where the samples can be analyzed using the Vertebrate Automated Screening Technology system with the Large Particle Sampler mounted onto a CLSM.
The Vertebrate Automated Screening Technology system with the Large Particle Sampler is a system that can either be mounted onto a CLSM or stereo microscope. This device allows the loading of live or fixed embryos from a 96-well plate or bulk container automatically through a glass capillary, and orientates it in front of the camera at the desired angle (e.g., dorsal or lateral). Images of the embryo in all orientations can be made with the on board camera or with an external CLSM (Figure 7). Embryos will subsequently be transferred in the collection or waste container.
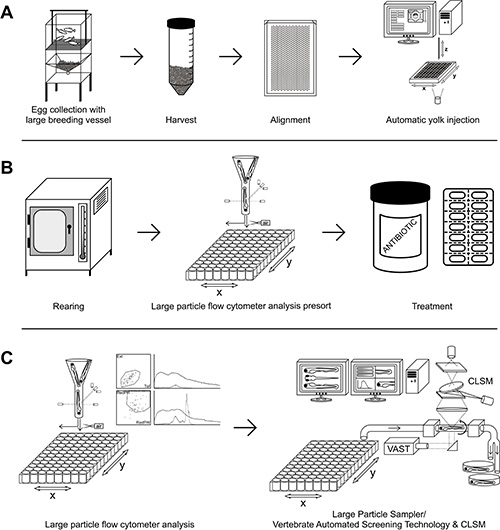
Figure 1. Mainstream experimental outline. A) Adult fish are put together to mate, eggs are collected, aligned into an agarose plate and injected. B) The injected eggs are incubated at 28 °C and will be pre-sorted for possible drug treatment. C) Subsequent analysis by large particle flow cytometer and/or Large Particle Sampler/Vertebrate Automated Screening Technology with CLSM. Please click here to view a larger version of this figure.
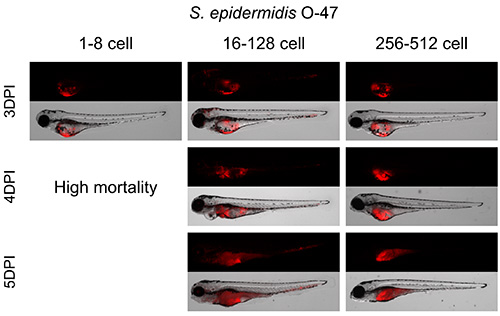
Figure 2. Establishment of best cell stage for S. epidermidis yolk injection. Zebrafish embryos were injected in the yolk at different developmental stages from 1 to 512 cell stage with 100 cfu of S. epidermidis. Embryos injected between 1 and 8 cell stage showed bacterial growth in the yolk and high mortality from 4 dpi. Embryos injected between 16 and 128 cell stage showed bacterial growth in the yolk and inside the body starting at 3 dpi. Embryos injected between 256 and 512 cell stage showed many bacterial growth inside the yolk. Please click here to view a larger version of this figure.
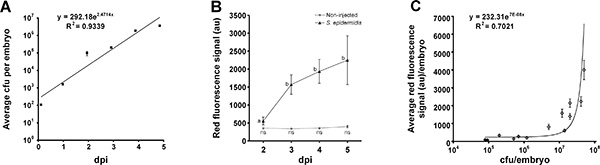
Figure 3. Quantification of bacterial burden using large particle flow cytometer. 100 cfu of S. epidermidis were injected into the yolk of zebrafish embryos. A) Up to 5 dpi, each day, groups of 10 embryos were homogenized and plated directly, showing the average exponential growth based on two biological replicas (error bars = SEM). B) Large particle flow cytometer analysis shows the average fluorescence signal from non-injected and S. epidermidis injected embryos. 30-160 embryos per condition were analyzed (error bars = SEM), different letters indicate statistical significant differences by one way ANOVA followed by Tukey's post-hoc test (P < 0.001), ns: not significant differences. C) Correlation between cfu and average fluorescence signal of groups of 10 S. epidermidis infected embryos (error bars = SEM). This figure has been modified from Veneman et al. (2013)8. Please click here to view a larger version of this figure.
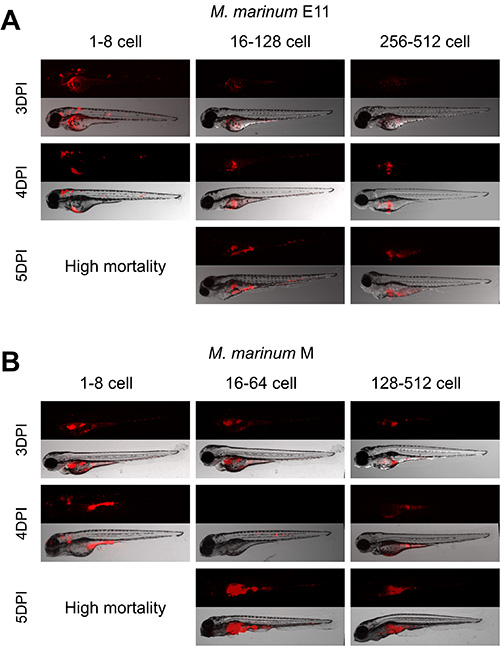
Figure 4. Establishment of the best cell stage for M. marinum yolk injection. Zebrafish embryos were injected at all the different developmental stages from 1 to 512 cell stage with 30 cfu of M. marinum E11 and M strain. A, B) Embryos injected from 1-8 cell stage showed similar spreading and mortality with both strains. A) Embryos injected between 16-128 cell stage with E11 strain showed formation of granulomas and systemic infection while those injected from 256 to 512 cell stage kept bacterial burden into the yolk. B) Embryos injected between 16-64 cell stage with M strain showed formation of granuloma like structures and systemic infection while those injected from 128 to 512 cell stage kept bacterial burden into the yolk. Please click here to view a larger version of this figure.
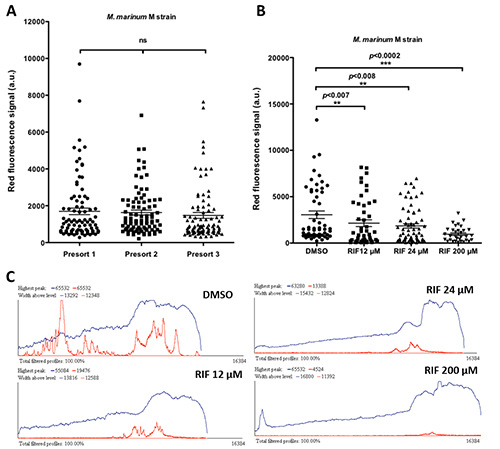
Figure 5. Treatment of M. marinum acute infection with a first-line anti-tuberculosis drug. Embryos injected between 16-64 cell stage with 30 cfu of M. marinum M strain were run through the large particle flow cytometer at 3 dpi to be sorted in two groups after discarding the non- and/or highly infected embryos. A) Fluorescence of individual embryos in both groups. B) Embryos treated with Rifampicin (RIF) for 48 hr at doses of 12, 24, and 200 μM were analyzed at 4 dpi; their bacterial load is significantly reduced. C) Representative COPAS profiles of embryos treated with DMSO and Rifampicin at doses of 12, 24, and 200 μM for 24 hr. Bacterial load and distribution is indicated by the red peaks. Blue line represents the profile of the element sorted (4 dpf zebrafish embryo) by the COPAS. 60-90 embryos per condition were analyzed. Each data point represents an individual embryo. Values are indicated as mean ± SEM. ns: not significant differences. Analysis of statistical significance of differences was performed by one way ANOVA followed by Tukey's posthoc test. Please click here to view a larger version of this figure.
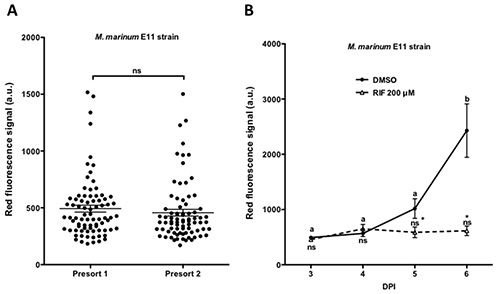
Figure 6. Treatment of M. marinum chronic infection with a first line antituberculosis drug. Embryos injected between 16-64 cell stage with 30 cfu of M. marinum E11 strain were run through the large particle flow cytometer at 3 dpi to be sorted in two groups after discarding the non- and/or highly infected embryos. A) Fluorescence of individual embryos in both groups. B) Embryos treated with Rifampicin (RIF) at 200 μM during 4 days were analyzed showing a significant reduction of the bacterial load after 1 day of treatment. 90 embryos per condition were analyzed. Values are indicated as mean ± SEM. Different letters indicate significant differences between time points of the same treatment. * indicates significant differences with control group. ns: not significant differences. Analysis of statistical significance of differences was performed by one way ANOVA followed by Tukey's posthoc test. (P < 0.05). Figure B) has been modified from Spaink et al. (2013)13. Please click here to view a larger version of this figure.
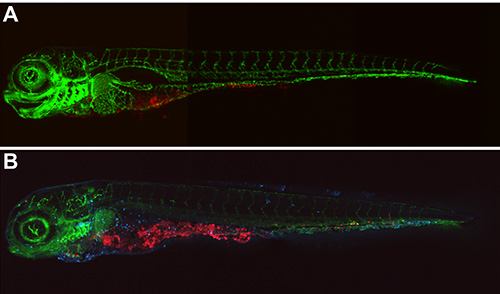
Figure 7. Result of M. marinum E11 yolk injection imaged using Vertebrate Automated Screening Technology and CLSM. Confocal Z stack (stitched 3 images) of a 5 dpi fli1-egfp14 embryo. A) Live embryo showing proliferation of M. marinum E11 bacteria (red) throughout the body. B) Fixed 5 dpi fli-egfp embryo showing M. marinum E11 bacteria (red) throughout the body co-localizing with leucocytes (light blue) detected by L-plastin immunostaining15.
