Sprague Dawley rat (Charles River) pups of both genders are used. Litters are culled to 12 on PD1 (birth being PD0). The dams are maintained on a 12 hr light/dark cycle with ad libitum access to food and water. Experimental procedures have been approved by Memorial University’s Institutional Animal Care Committee.
1. Nose Plug Construction
NOTE: This procedure was adapted and modified from Cummings et al. (1997)35.
- Aquire polyethylene-20 tubing and 3-0 silk suture thread.
- Cut a small piece of polyethylene-20 tubing to approximately 0.8 mm.
- Thread silk suture through the prepared tubing such that there is thread on either side of the section of tubing.
- On one end of the thread outside of the plug, tie a knot in the thread.
- Pull the section of tubing down over the knot in the thread. The knot should lodge inside the tubing.
- Trim both ends of the thread such that ~2 mm of thread is protruding from one end of the tubing (see Figure 1A).
2. Naris Occlusion before Training
- Remove pup from dam and place in a secure dish covered with regular bedding.
- Use a cotton tip application to dab a local anesthetic jelly, 2% Xylocaine, on the naris to be occluded.
- Allow the pup to rest in the dish for ~3 min.
- Hold the pup gently but securely in the non-dominant hand.
- Using the dominant hand, pick up a nose plug and dab the same local anesthetic jelly around the tip from which the thread is not protruding. This will act as both an anesthetic for any minor pains associated with plug insertion and will act as a lubricant inside the naris. NOTE: The effect of Xylocaine starts within a couple of minutes and lasts 20-30 min. Pups in general show good tolerance for plug insertion after xylocaine jelly application (minimum struggling and vocalization).
- Gently insert the nose plug by firmly holding the pup and slowly rotating the plug with very gentle pushes until the plug is fully inserted and only the 2 mm thread is protruding from the naris (see Figure 1B). There should be no bleeding from either naris during this process. Pups with bleeding during nose plug insertion are excluded and returned to dams
- Allow the animal to rest in this dish for 5 min in order to habituate to the plug.
- Remove the pup from the habituation dish and begin the conditioning paradigm24.
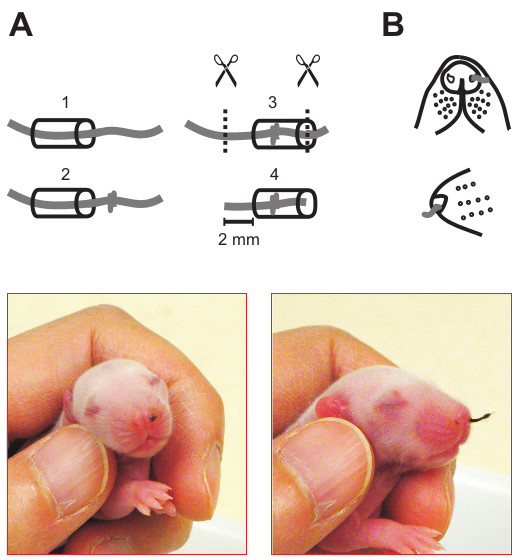
Figure 1. Construction of a nose plug. A) Schematics showing the steps of making a nose plug. A thread is pulled through polyethylene tubing; a knot is made and pulled into the middle of the tubing to block it; two ends are cut with a 2 mm residue in one end out of the tubing. B) Front and lateral view of a rat with a nose plug in one naris.
3. Scented Bedding Preparation
- Wearing new gloves and in a fume hood to prevent odor contamination, place 500 ml of woodchip bedding into a plastic bag.
- Use a syringe to draw up 0.3 ml peppermint extract, and spray this over the bedding in the plastic bag.
- Tie the bag shut, shake the bag vigorously, and allow the bedding to rest in the bag for 5 min.
- Place the scented bedding in a clear, shallow, acrylic training box (20 x 20 x 5 cm3, Figure 2A) uncovered in a fume hood for 5 min before use. Once the bedding is prepared, discard these gloves, and do not allow these gloves to come in contact with the animals.
- Place the unscented bedding in an identical clear plastic box, and ensure that it does not come into contact with the scented bedding or used gloves.
4. Odor Conditioning Paradigm (See Picture in Figure 2A)
Pups undergo either a single conditioning session, on PD 6, or multiple trial sessions (one session per day, PD 3-6).
- Place the habituated pup on scented bedding. For control odor only (O/S–) pups, leave these pups on the bedding for 10 min, then skip to step 4.5. For experimental odor + stroke (O/S+) pups, continue to the following steps in this section.
- Stroke the pup for 30 sec using a small paintbrush. Use rapid circular motions primarily around the hind region of the pup.
- Allow the pup to rest for 30 sec.
- Repeat steps 4.2 & 4.3 for a total of 10 min (i.e. 10 pairings of stroking + odor).
- Remove pup from the conditioning box, remove the nose plug and return the pup to the dam.
5. Lateralized Odor Preference Testing (See Picture in Figure 2B)
Testing occurs at various time points (e.g., 24 or 48 hr) following the final training session. Testing is carried out in a stainless steel testing chamber (30 x 20 x 18 cm3), which is placed on top of two training boxes (training box is described in 3.4), separated by a 2 cm neutral zone. One training box contains peppermint-scented bedding while the other box contains clean, unscented bedding. The floor of the testing chamber is a metal grid, which is then covered by a removable sheet of plastic mesh (Figure 2B).
- Prepare one peppermint and one unscented bedding as per Section 3, and place each box under opposite sides of the testing chamber, 2 cm apart. Place the plastic mesh on the metal-grid floor of the testing chamber.
- Remove the pup from the dam and place a firm dab of odorless silicone grease on the naris that is occluded during training. Re-apply the grease throughout the first testing procedure as needed. NOTE: Random naris occlusion during training and testing may be considered to avoid bias.
- Place pup in the neutral zone of the testing chamber.
- Allow the pup to explore the chamber for 1 min, recording how long the pup spent over the two sides of the chamber (i.e. over peppermint or neutrally-scented bedding).
- Allow the pup to rest for 1 min in a covered plastic holding chamber.
- Repeat steps 5.2 & 5.3 for a total of 10 min (i.e. 5 test trials separated by 5 rest trials) switching the initial orientation of the pup in the chamber in order to control for direction preferences.
- Immediately following testing, wipe away the grease from the naris.
- Insert a polyethylene noseplug into the opposite naris as per section 2 and allow the animal to rest for 10 min.
- Test the pup once again as in 5.3 -5.6, remove the plug, and return the pup to the dam. Remove and clean the plastic mesh of the testing chamber with 95% ethanol and allow the liquid to evaporate. Place the mesh back before testing the next pup.
NOTE: Applying silicone grease at the first naris occlusion during testing prevents the chance of bleeding and stress associated with nose plug insertion.

Figure 2. Early odor preference training and testing. A) Early odor preference training using odor + stroking paradigm. B) Two choice odor preference testing with peppermint bedding on one side, control unscented bedding on the opposite side. A 2 cm neutral zone is placed in between.
6. Testing the Effectiveness of Single Naris Occlusion
This experiment is performed to determine whether single naris occlusion leads to lateralized activation of the olfactory system.
- Perform unilateral naris occlusions on PD 6 or 7 pups as described in section 2.
- After ~5 min habituation, place the pup in a covered plastic container and expose it to 30 µl pure peppermint oil soaked in a piece of tissue for 10 min.
- Immediately after the peppermint odor exposure, inject the pup intraperitoneally (i.p.) with chloral hydrate (400 mg/kg) as a general anesthetic, or pentobarbitol, (150 mg/kg).
- Once fully anesthetized (showing no response to tail or foot pinch), transcardially perfuse the pup with ice-cold solutions of saline (0.9%) for ~1 min, followed by paraformaldehyde (4%, dissolved in 0.1 M phosphate buffer solution, PBS).
- After 10 min of paraformaldehyde perfusion, collect the brain and place it in paraformaldehyde overnight at 4 °C, then transfer the brain to a sucrose solution (20% in PBS) for an additional 24 hr.
- Cut coronal brain slices at 30 μm thickness with a cryostat. Collect OB and PC slices and mount onto gelatin-coated slides, followed by standard immunohistochemistry staining for pCREB antibody21,25,30.
7. Testing the Reversibility of Single Naris Occlusion
This experiment tests whether the blocking effect is reversible at 24 hr following the removal of the nose plug.
- Perform unilateral naris occlusions on PD 6 or 7 pups as described in section 2.
- After 15 min (equivalent to the duration of the naris occlusion during training – 5 min habituation + 10 min training), remove the nose plug, and return the pup to the dam.
- 24 hr later, expose the pup to the peppermint odor in a covered plastic container for 10 min as described in 6.2.
- Follow same steps in sections 6.3-6.6.
Here we review some of the previously established results24 to demonstrate the effectiveness of the naris occlusion in isolating odor input and learning to one hemisphere, and the reversibility of this method.
Single naris occlusion during early odor preference training leads to a lateralized odor memory24. The memory is confined to the spared naris (Figure 3). When pups are tested for odor preference with the same naris occluded as during training, they show preference for the conditioned odor (e.g., peppermint). When pups are tested with the opposite naris occluded, they show no preference for the conditioned odor. Together, these results suggest that an odor preference memory is only formed and expressed through the spared naris that underwent the odor + stroking associative conditioning.
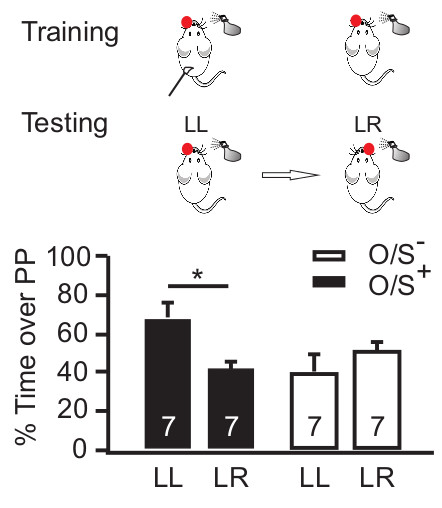
Figure 3. Single naris occlusion induces lateralized odor learning. The behavioral protocol is shown in the upper panel. Odor + stroking (O/S+) or odor only (O/S–) animals with single naris occluded during training, underwent odor preference testing first with the same naris occluded then with the opposite naris occluded. The lower panel shows the percentage of time spent over peppermint-scented bedding among different groups in a two choice odor test. *p < 0.05. Error bars, mean ± SEM. Reproduced from Fontaine et al. J. Neuroscience (2013) with permission.
The lateralized odor training results in lateralized activation of the olfactory system during odor exposure (Figure 4)24. Single naris occlusion prevents activation of the OB and the PC of the ipsilateral hemisphere during odor exposure. This is demonstrated by monitoring CREB phosphorylation in the OB, and the PC. As shown in Figure 4, using immunohistochemistry, phosphorylated CREB (pCREB) is significantly less in the occluded hemisphere following peppermint odor exposure, compared to the contralateral spared hemisphere. Nissl staining (Figure 4A) demonstrates comparable cell bodies in the mitral cell layer of the OB, and in the pyramidal cell layer of the PC of both hemispheres. However, pCREB is significantly less in both cell layers in the hemisphere ipsilateral to the occluded naris (Figure 4B).
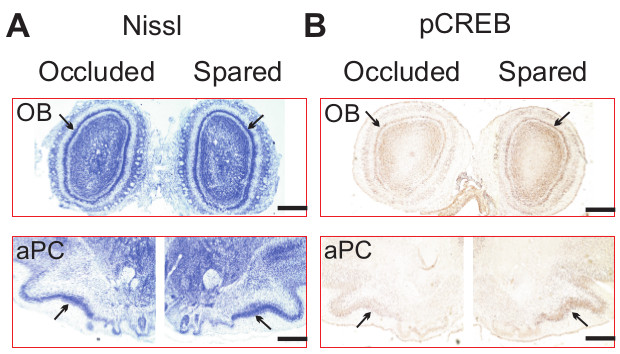
Figure 4. Single naris occlusion results in lateralized activation of the olfactory system during odor exposure. A) Nissl staining of the olfactory bulb (OB) and anterior piriform cortex (aPC). B) PCREB expressions in the occluded and spared hemispheres following peppermint exposure in a single naris occluded pup. Arrows indicate mitral cell layer in the OB and pyramidal cell layer in the aPC. Scale bars, 500 μm. Reproduced from Fontaine et al. J. Neuroscience (2013) with permission.
The effect of a single trial (15-20 min) naris occlusion is transient and reversible, and does not result in visible longer-term neuronal damage that could lead to altered odor perception and reduced neuronal activation to odors during testing24. As indexed by pCREB staining in the OB and the PC (Figure 5), pCREB expressions in mitral cells of the OB, and the pyramidal cells in the PC to odor exposure are comparable between the occluded and spared hemispheres, 24 hr following the removal of the nose plug – the same time point that odor preference testing is carried out following early odor preference training.
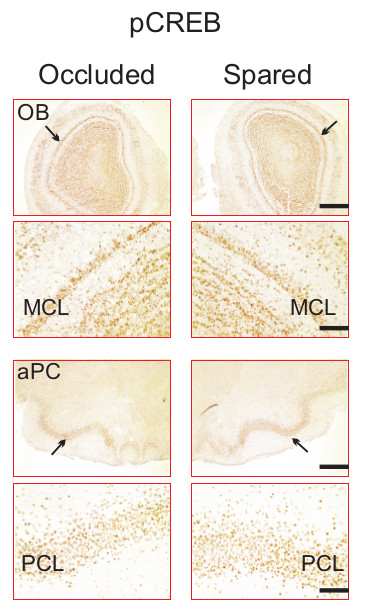
Figure 5. Assessment of neuronal reactivity following reversible naris occlusion. PCREB staining of the OB and the aPC, 24 hr following the removal of a nose plug in one pup. Arrows indicate mitral cell layer in the OB and pyramidal cell layer in the aPC. MCL, mitral cell layer. PCL, pyramidal cell layer. Scale bars, 500 µm for lower magnification and 100 μm for higher magnification.
