Because animals undergoing this procedure do not require specialized post-operative care, short or long-term time survival periods are easily incorporated into the study, depending on the need to investigate acute or chronic pathology following injury. Principal features of reactive gliosis, such as upregulation of GFAP and hypertrophy of soma, can be observed as early as 2-3 days following injury. The peak phase of proliferation for reactive astrocytes is during days 3-5 following injury10. The representative results shown below are from animals that received a stab wound lesion 7 days earlier.
The general morphology and cytoarchitecture of the forebrain following a forebrain stab injury can be visualized by Nissl staining (Figure 2). Although the blade track is most prominent throughout the center of the lesion, the disrupted cortical cytoarchitecture reveals the rostral and caudal extent of the damaged tissue. Reactive astrocytes can be observed by immunohistochemistry for GFAP (Figure 3). Note that many cortical astrocytes do not exhibit immunohistochemically detectable levels of GFAP in the absence of injury. However, GFAP expression is dramatically upregulated in the hemisphere ipsilateral to the injury while remaining at relatively low levels in the contralateral hemisphere (Figure 3), suggesting that reactive astrocytes are restricted to the ipsilateral hemisphere. Note that while the cortical tissue ipsilateral to the lesion demonstrates marked upregulation of GFAP, other astrocytic markers, such as S100β are constitutively expressed in the absence of an injury (Figure 4), and maintain similar expression levels following injury (Figure 4). In addition to increased GFAP expression, reactive astrocytes undergo cellular hypertrophy. Cell bodies and processes become enlarged and show intense staining for GFAP (Figure 3).
The proliferation of reactive astrocytes can be observed by administering the thymidine analog, 5-bromo-2'-deoxyuridine (BrdU), or by immunostaining for the proliferative markers Ki67 or PCNA. We routinely administer 200 mg/kg BrdU, i.p., to animals during days 3-5 following injury, the peak of reactive gliosis10 (Figure 3). However the precise dosage and timing of BrdU should be independently considered for each study, bearing in mind that BrdU will permanently label cells undergoing proliferation, as well as their progeny, at the time of administration, but that cells that enter the cell cycle before BrdU starts, or after BrdU administration is complete, will not be marked. In Figure 4, we show extensive co-localization between GFAP and BrdU at 1 week post injury, indicating that many reactive astrocytes proliferated during the time course of BrdU administration. Note that proliferating reactive astrocytes are predominantly localized adjacent to the lesion core, whereas reactive astrocytes localized distal from the lesion core are largely non-proliferative (Figure 4).
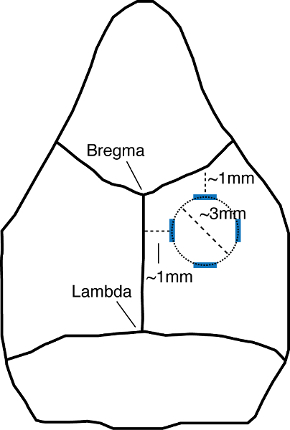
Figure 1: Schematic of the mouse skull, depicting the area of the craniotomy. Blue lines depict the initial markings identifying the boundaries of the area to be drilled. The top and left marks are measured at 1 mm below or lateral to the coronal or sagittal sutures, respectively. The bottom and right marks are measured at 4 mm from the coronal and sagittal sutures, respectively. The craniotomy is performed by drilling a circle within the marked boundaries (dotted line), creating a craniotomy that is roughly 3 mm in diameter (dashed line). Schematic is not to scale.
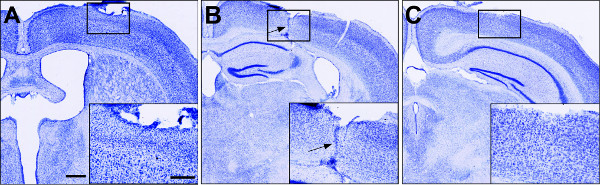
Figure 2: Nissl staining throughout the rostral-caudal extent of the lesion volume. (A–C) Coronal slices of injured brains 1 week post injury, showing the hemisphere ipsilateral to the lesion. Insets depict zoomed in images of the boxed regions. The blade track is most prominent in the center of the lesion, ~2.5 mm from Bregma (arrow in B). Note the disrupted cortical cytoarchitecture (insets) in sections anterior (A) and posterior (C) to the lesion center. Scale bar 500 μm, inset, 250 μm. Please click here to view a larger version of this figure.
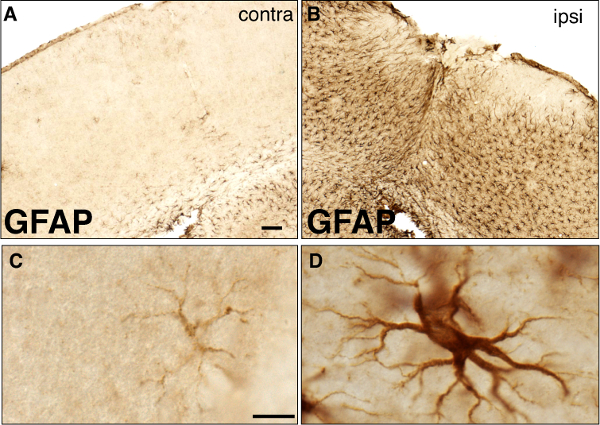
Figure 3: Brightfield immunohistochemistry for GFAP 1 week following a forebrain stab injury. (A–B) Low magnification images of GFAP staining in the contralateral (A) and ipsilateral (B) hemispheres of the same tissue section from an injured animal. The lesion site is shown in (B), and the corresponding region in the uninjured contralateral hemisphere is shown in (A). Scale bar, 100 μm. (C–D) High magnification images of normal (C) and reactive (D) astrocytes from the contralateral and ipsilateral hemispheres, respectively. Note the dramatic hypertrophy of the cell body and processes of reactive astrocyte in (D), compared to (C). Scale bar, 10 μm.
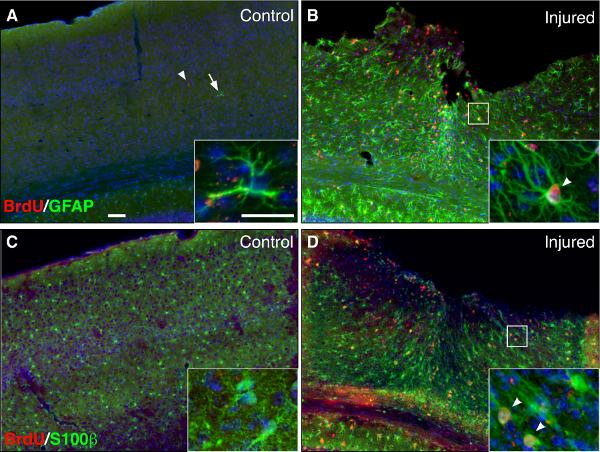
Figure 4: Reactive astrocytes proliferate following a forebrain stab injury. (A–B) Immunofluorescent staining for BrdU (red) and GFAP (green) in uninjured, control (A) and injured (B) brains, 1 week following stab injury. (C–D) Immunofluroescent staining for BrdU (red) and the astrocytic marker S100β (green) in the uninjured (C) and injured (D) hemispheres, 1 week following stab injury. Animals received BrdU over days 3-5 post injury. Note that many astrocytes are proliferating at the lesion site in the injured cortex (B and D, insets, arrowheads), whereas astrocytes in the uninjured cortex are not proliferating (A and C, insets). Counterstaining with DAPI (blue). Scale bars, 100 μm, inset, 25 μm.
