Microscopy analysis of fission yeast sexual reproduction
1. Media Preparation
- Prepare Minimum Sporulation medium (MSL-N)15 by mixing the following components: Glucose: 10 g/L, KH2PO4: 1 g/L, NaCl: 0.1 g/L, MgSO4·7H2O: 0.2 g/L. Add Trace elements (10,000x): 100 µl/L, Vitamins (1,000x): 1 ml/L, and 0.1 M CaCl2 ml/L. Filter-sterilize using a 0.22 μm pore size filter and store at room temperature (RT).
- Use the following Vitamins (1,000x) stock: Pantothenate: 1 g/L, Nicotinic Acid: 10 g/L, Inositol: 10 g/L, Biotin: 10 mg/L. Filter-sterilize using a 0.22 μm pore size filter and store at 4 °C.
- Use the following Trace elements (10,000x) stock: Boric Acid: 5 g/L, MnSO4: 4 g/L, ZnSO4·7H2O: 4 g/L, FeCl2·6H2O: 2 g/L, MoO3: 0.4 g/L, KI: 1 g/L, CuSO4·5H2O: 0.4 g/L, Citric Acid: 10 g/L. Filter-sterilize using a 0.22 μm pore size filter and store at 4 °C.
- Prepare MSL-N Agarose 2% (used to make agarose pad chambers) by combining 10 ml of MSL-N with 0.2 g of agarose. Melt for ~2 min at high power in a microwave oven until agarose dissolves and aliquot 0.5 ml in microcentrifuge tubes. Store at RT.
- Prepare Minimum Sporulation medium with nitrogen (MSL+N) from MSL-N by adding (NH4)2SO4: 2 g/L, Leucine: 0.225 g/L, Adenine: 0.225 g/L, Uracil: 0.225 g/L. Filter-sterilize using a 0.22 µm pore size filter. Store at RT.
Note: Leucine, adenine and uracil are added to this medium to allow the growth of auxotrophic strains. Additional amino-acid supplement should be included if the strain used carries a distinct auxotrophy (for instance his3Δ). Please also note that work with fully prototroph strains is recommended, as only such strains will allow high mating efficiency. - Prepare 200 ml VALAP for chamber sealing. In a beaker add equal weights of Lanolin, Vaseline (or other petroleum jelly), and Paraffin. Heat mixture at low temperature and stir occasionally until thoroughly blended. Aliquot the mix into several small petri dishes. Store at RT.
2. Culturing Fission Yeast Strains for Mating Experiments (Figure 1).
- Day 1, Evening:
Inoculate freshly streaked strains from solid media into culture tubes containing 3 ml of MSL+N. Use flat-bottom tubes of about 2.5 cm diameter. Use other culture tubes or flasks with adapted culture volume. Incubate overnight (O/N) with shaking at 25 °C, 200 rpm. If working with heterothallic strains, inoculate separately.- Dilute cell suspensions in media the following morning to ensure that cultures have optical density measured at 600 nm (O.D.600) of 0.4-0.8 in the evening.
Note: O.D.600 measurements as a proxy of cell concentration for fission yeast are linear in the range of approximately 0.1-1.0. For our spectrophotometer O.D.600 = 0.1 corresponds to 1.4 x 106 cells per ml. Thus, if initial O.D.600 reads are outside this range samples need to be diluted/concentrated for reliable measurements.
- Dilute cell suspensions in media the following morning to ensure that cultures have optical density measured at 600 nm (O.D.600) of 0.4-0.8 in the evening.
- Day 2, Evening:
Dilute cells in 20 ml of MSL+N media to O.D.600 = 0.025 in 100 ml flasks. Incubate strains O/N at 30 °C, 200 rpm.
Note: Wild-type cell cultures should reach O.D.600 ~0.8 the following morning (15 hr later). If working with strains with longer generation time adjust dilution accordingly. - Day 3, Morning:
Measure O.D.600 to verify that cultures have cell density of O.D.600 ~0.8. If working with heterothallic strains, mix equal numbers of partner cells. Pellet cells at 1,000 x g and transfer to a 1.5 ml tube. Wash cells three times in 1 ml of MSL-N medium. Re-suspend cells in 3 ml of MSL-N medium and dilute cells to O.D.600 = 1.5.- To monitor mating between sister cells directly mount cells for imaging (see Protocol Section 2.4). Use homothallic h90 strains, in which mating type switching will take place during the mitotic divisions occurring after nitrogen deprivation. Immediate mounting of cells on the agarose pad ensures that, after cell division, sister-cells remain next to each other.
- To monitor cell polarization (exploratory dynamics and shmooing) incubate 1-3 ml of cell culture at 30 °C, 200 rpm for 3-4 hr prior to mounting cells for imaging. Within these 3-4 hr, the last mitotic division will have occurred. A few cells will have initiated polarization, but most will just be starting their exploratory polarization dynamics.
- To monitor cell-cell fusion incubate 1-3 ml of cell culture at 30 °C, 200 rpm for 4-6 hr prior to mounting cells for imaging.
- To monitor post-fusion events incubate 1-3 ml of cell cultures at 30 °C, 200 rpm for 8 hr prior to mounting cells for imaging.
- To monitor response to a specific pheromone concentration (only for heterothallic strains) incubate 3 ml of cell culture at 30 °C, 200 rpm for 3-4 hr prior to mounting cells for imaging.
- Remove the MSL-N-containing microcentrifuge tube from the 95 °C heat block (see Protocol section 2.4.1. below). Add P-factor or M-factor at the desired concentration directly in the melted MSL-N agarose. Mix well by vortexing before immediate pad preparation.
- After cell mounting, incubate the pad for 15-30 min at 25 °C prior to imaging. P-factor and M-factor pheromones were used from a stock solution of 1 mg/ml in methanol.
Note: If working with mutant strains incubation times may vary. Clumping of cells may occur after prolonged incubation in liquid media and may be particularly strong in some mutant strains. Clumped cells cannot be spotted onto agarose pads in a single layer and thus are difficult to autofocus and image. In case of extensive cell clumping mount cells on agarose pads immediately after washes and re-suspension in nitrogen-lacking media (as in the Protocol Section 2.3.1.) and incubate them at 30 °C prior to imaging.
- Mounting Cells for Imaging:
- Prepare MSL-N agarose pads16 by melting an aliquot in a 95 °C heat-block for 10-15 min and adding 200 μl between two glass slides separated by spacers (Figure 1B). Use spacers with a thickness of ~0.5 mm. To make spacers, use strips cut out of cardboard, such as that delivered with restriction enzymes.
- Pellet 100 μl of cells at 1,000 x g for 1 min, remove the supernatant and re-suspend cells in residual 2-4 μl medium. Do NOT re-suspend in fresh medium as it delays mating.
- After 2-3 min carefully remove spacers and the top glass slide. If several strains are to be mounted, prepare the cell concentrates before preparing the agarose pad.
- Add 1 μl of cells to the pad and wait until the drop starts drying (~1 min) before covering with cover slip and sealing with VALAP.
- Let the pad sit for at least 30 min before imaging at 25 °C or RT.
- To obtain a mixture of cells in various stages of mating, make a pad immediately after washes in MSL-N (see Section 2.3.) and incubate at 18 °C O/N (~15 hr). This approach is useful for imaging cells at distinct mating stages with high temporal resolution or samples with weak signal.
3. Live-cell Imaging of Mating Yeast Cells
- Adjust Image Acquisition Settings.
Note: Image acquisition settings presented here were optimized for the DeltaVision platform composed of a customized Olympus IX-71 inverted microscope with Plan Apo 60X/1.42 NA or U-Plan Apo 100X/1.4 NA objectives, a CoolSNAP HQ2 camera and an Insight SSI 7 color combined unit illuminator. The hardware is controlled by softWoRx v4.1.2 that also enables digital autofocusing.- Ensure that the autofocusing intervals do not exceed 15 min since the Z-drift over longer time can surpass the capacity of the software autofocusing system. If using a hardware-based autofocusing system, use larger intervals.
- Adjust imaging interval. Take images every 10 min for ~15 hr as a starting point when working with a fluorescently tagged protein for the first time. This interval provides a good overview of the entire mating process without extensive bleaching.
- Image with 5 min intervals or shorter to more precisely time events such as fusion. Shorter intervals require strong fluorophores enabling low exposure times and little bleaching. For imaging with intervals below 1 min avoid O/N microscopy and instead prepare a mixture of all mating stages as detailed in Protocol Section 2.4.6.
- Adjust image exposure time. Test exposure times between 50 and 300 msec. Aim to strike a balance between the signal-to-noise ratio and photobleaching as to achieve a large number of time-points with a discernable signal (typically over 100).
- Adjust Z-sectioning. Sections every 0.3 µm covering 5 µm provide a good imaging depth. Note that Z-sectioning further increases photo-damage, which can be minimized by using the OAI (optical axis integration – a single sweep acquisition of the total sample depth). Good auto-focus system reduces the need for many Z-sections and is highly recommended.
- Tips to Study Mating Type-specific Behaviors:
- For heterothallic strains, use h- and h+ cells expressing distinct fluorophores, so that the two cell types can be distinguished. Use any genetically encoded fluorophore expressed as a cytosolic version or fusion to an endogenous protein, though we have had good success with sfGFP, a fast-folding variant of GFP17. Endogenous proteins are usually tagged at their endogenous genomic locus by homologous recombination14.
- For homothallic strains, express a genetically encoded cytosolic fluorescent protein under the control of mating type specific promoters such as those of the pheromone receptor genes map3 and mam2 to drive the expression in P and M cells, respectively. The promoters of map3 and mam2 are inactive during vegetative growth and are induced upon nitrogen starvation18,19. Constructs driving the expression of cytosolic GFP or mCherry under the control of these promoters are typically integrated at a genomic locus (ura4, leu1) that does not interfere with the normal progression of the sexual life cycle12.
- Use mating type specific cytosolic fluorophores (as in Protocol Section 3.2.2) to determine precise timing of cell fusion visualized as the transfer of fluorescent signal from one partner cell to the other.
4. Quantification of Mating and Fusion Efficiencies
- To quantify mating and fusion efficiencies11,12, spot cells directly after MSL-N wash onto MSL-N agarose pads, as in step 2.3.1 above, and leave for 24 hr at 25 °C prior to imaging (Figure 1A). Using this protocol a homothallic wild-type strain mating efficiency reaches ~60% and fusion efficiency 99%. To test whether any reduction in these values observed in mutant strains reflect a terminal phenotype or a delay, repeat the experiment with a 36 hr incubation time.
- Calculate mating efficiency as
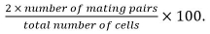 Mating pairs include zygotes, asci, and unfused cell pairs identified from their position and growth towards each others.
Mating pairs include zygotes, asci, and unfused cell pairs identified from their position and growth towards each others. - Calculate fusion efficiency as
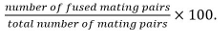
Note: Fused mating pairs are identified by their ability to form spores if it is known that the mutant under study does not influence sporulation. If sporulation cannot be used as read-out, fusion is determined by observing the spreading of a cytosolic marker expressed in only one mating partner into both partners.
Fission Yeast Growth and Mating Dynamics Upon Removal of a Nitrogen Source
As nitrogen starvation is a prerequisite for initiation of sexual reproduction in fission yeast, wild-type homothallic h90 strain was monitored upon shift from nitrogen-rich to nitrogen-deprived medium (Figure 2), following the protocol outlined in Figure 1. Briefly, cells were grown O/N to exponential phase (O.D.600 = 0.5) in MSL+N medium, collected, washed and re-suspended in MSL-N liquid medium to final O.D.600 = 1.5. Every 2 hr aliquots of cells were calcofluor-stained and imaged (Figure 2A–D).
Calcofluor staining (Figure 2A) revealed that in nitrogen-rich medium ~21% of cells were septating (n >300, Figure 2B) and that average cell length at division was 15.2 ± 1.4 µm (n = 50, Figure 2C). Non-dividing cells showed a broad distribution of cell lengths (8-14 µm; n >200, Figure 2D). However, two hours after the shift to MSL-N medium there was a drastic change in the length distribution of non-dividing cells, with over 60% of cells being shorter than 9 µm (n >200, Figure 2D). Cell septation was also detected at the reduced length of 9.8 ± 0.8 µm (n = 50, Figure 2A, 2C). A further reduction in length of both non-dividing and dividing cells was also observed, as time in MSL-N increased. Indeed, after 8 hr the septation index of the culture decreased below 2% (n >300) and over 85% of cells arrested their cell cycle at lengths shorter than 7 µm (n >200, Figure 2D).
Cells started to form mating pairs four hours after the shift to liquid MSL-N medium (<1%, Figure 2A). Subsequently the number of mating pairs rapidly increased and 8 hr after the shift 10% of cells engaged a mating partner (n >200, Figure 2B).
To monitor mating dynamics, 6 hr after starvation induction, an aliquot of cells was also mounted onto MSL-N agarose pad for imaging. Cells underwent shmooing, fusion and sporulation (Figure 2E, 2F) in the subsequent 21 hr without any apparent synchrony (Movie S1). The mating efficiency depended on the relative position and density of cells in a given field of view, with over 60% of cells engaged a partner when positioned densely in a monolayer (Figure 2G and Movie S1).
The wild-type heterothallic h+ and h- fission yeast strains were also subjected to the same protocol with an additional step of mixing h+ and h- cells in a one-to-one ratio at the time of nitrogen removal. Similar starvation and mating dynamics were observed, but the mating efficiency was slightly lower (Figure 2G, Movie S2).
Monitoring Dynamics of Fluorescently Tagged Proteins in Mating Fission Yeast Cells
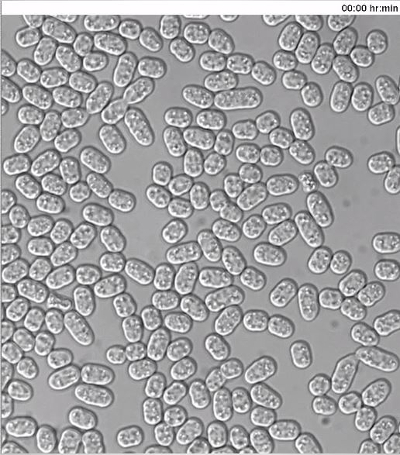
To monitor the dynamics of type-V myosin Myo52 (Ref20) in mating cells, exponentially grown h- cells expressing Myo52-3GFP from the native locus were mixed with h+ cells similarly expressing Myo52-tdTomato, as well as cytosolic GFP from the P-cell specific map3 promoter. The cell suspension was washed, diluted in MSL-N medium and incubated for 6 hr at 30 ºC, 200 rpm prior to mounting cells onto MSL-N agarose pads for O/N imaging at 10 min intervals.
As previously reported11,12 Myo52 initially formed dynamic zones throughout the cell cortex (Figure 3A, arrowheads and Movie S3). Myo52 signal was then stabilized (Figure 3A, arrows and Movie S3) in a single focus just before the fusion event, which was visualized by the transfer of the cytosolic GFP signal from the h+ into the h- cell (Figure 3A and Movie S3). The Myo52-tdTomato signal could also be observed at the fusion neck during its expansion but no discernable signal was evident as the zygote proceeded to sporulation (Figure 3A and Movie S3).
Monitoring the Behavior of Heterothallic Fission Yeast Cells Exposed to External Pheromones
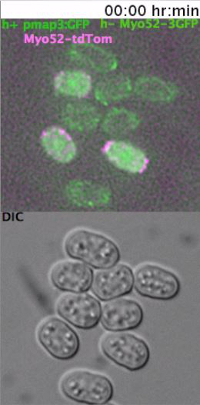
Heterothallic h- cells lacking the Sxa2 protease that degrades P-factor for desensitization readily respond to synthetic P-factor. An h-sxa2Δ strain, expressing Scd2-GFP as a marker for active Cdc4211, was grown in MSL+N medium, accordingly to the described protocol. Cells were washed in MSL-N medium and incubated for 4 hr at 30 ºC. MSL-N agarose pads containing methanol (data not shown), 0.1 μg/ml (low pheromone, Figure 3B) or 1 μg/ml (high pheromone, Figure 3C) of P-factor were prepared, cut and positioned on a new slide to generate mini-pads containing different pheromone amounts. 0.5 μl of cell suspension was mounted on each mini-pads for O/N imaging at 5 min intervals.
In concordance with published results11, low P-factor levels promoted the formation of dynamic Scd2 zones without growth (Figure 3B, Movie S4), while high levels of P-factor stabilized a single Scd2 zone, thus inducing shmoo elongation from one cell pole (Figure 3C, Movie S5).
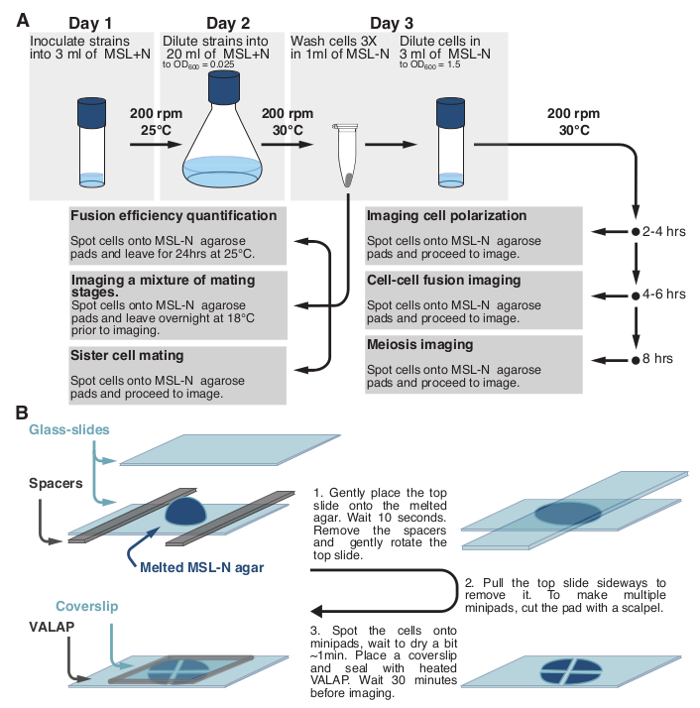
Figure 1: Schematic Representation of the Protocol. (A) Schematic description of the protocols used to monitor fission yeast sexual lifecycle and (B) to prepare fission yeast samples for long-term imaging. Please click here to view a larger version of this figure.
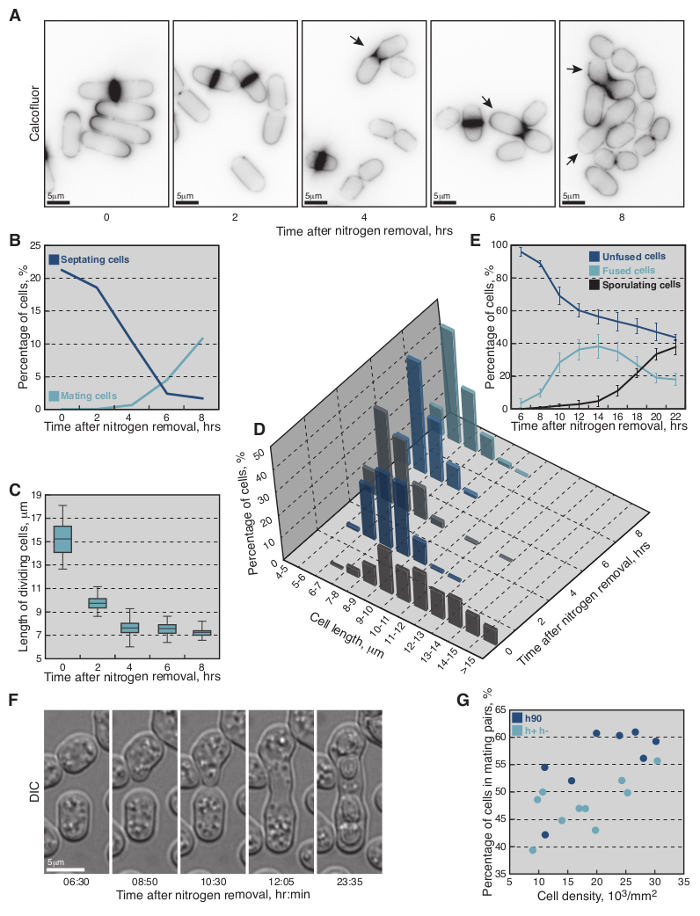
Figure 2: Growth and Mating Dynamics of Fission Yeast Cells Shifted from MSL+N to MSL-N Media. (A) Epifluorescence micrographs of calcofluor-stained cells shifted to MSL-N media for the indicated periods of time. (B) Percent of septating (dark-blue line, n >300 per timepoint) and mating (light-blue line, n >300 per timepoint) cells shifted to MSL-N media for the indicated periods of time. (C) Average length of septating cells shifted to MSL-N media for the indicated periods of time (n = 50 per timepoint except 8 hr timepoint where n = 20). (D) Length distribution of non-dividing cells shifted to MSL-N medium for the indicated periods of time (n >200 per timepoint). (E) Average fraction of unfused (dark-blue line), fused (light-blue line) and sporulating (black line) cells in a population of h90 wild-type yeast shifted to MSL-N media for the indicated periods of time and mounted onto an agar pad at timepoint 6 hr (n >900 cells per timepoint from three different timelapses). The cells were considered fused when no refractive cell wall between partners was visible in the DIC micrographs and formation of spore cell wall was used to score for sporulating cells. (F) DIC micrographs of mating cells shifted to MSL-N media for the indicated periods of time. (G) Mating efficiency as a function of cell density on agarose pads for h90 (dark-blue dots) and h+/h- (light-blue dots) cells shifted to MSL-N media for 24 hr. The most suitable density of cells for long-term imaging is achieved at approximately 25,000 cells/mm2. Please click here to view a larger version of this figure.
Figure 3: Dynamic Localization of Fluorescent Proteins during Fission Yeast Mating. (A) Deconvolved single z-plane epifluorescence micrographs of cells with the indicated genotypes shifted from MSL+N to MSL-N media for 6 hr and mounted onto MSL-N agarose pad for the indicated time. Arrowheads indicate dynamic Myo52-3GFP zones and the arrow points out Myo52 localization at the fusion focus. (B), (C) Deconvolved single z-plane epifluorescence micrographs of cells with indicated genotypes shifted from MSL+N to MSL-N media for 4 hr and mounted on MSL-N agarose mini-pads containing 0.1 μg/ml (B) or 1 μg/ml (C) of P-factor for the indicated time. Arrowheads indicate dynamic (B) or stable (C) Scd2-GFP zones. Please click here to view a larger version of this figure.
Supplemental Movie 1: DIC timelapse of wild-type h90 fission yeast cells that were nitrogen-starved for 6 hr prior to imaging. (Right click to download).

Supplemental Movie 2: DIC timelapse of wild-type h+ and h- fission yeast cells that were mixed and nitrogen-starved for 6 hr prior to imaging. (Right click to download).
Supplemental Movie 3: Deconvolved single z-plane epifluorescence and DIC timelapse of h- cells expressing Myo52-3GFP from the native locus mixed with h+ cells expressing Myo52-tdTomato from the native locus and cytosolic GFP from the P-cell specific map3 promoter. Cells were grown in MSL-N medium for 6 hr at 30 ºC prior to imaging. Transfer of cytosolic GFP into the h- cell defines the fusion time. (Right click to download).
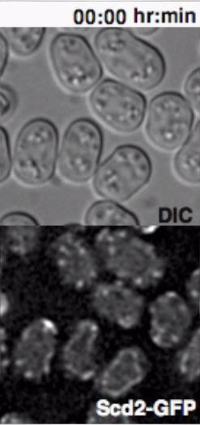
Supplemental Movie 4: Deconvolved single z-plane epifluorescence and DIC timelapse of h-sxa2 cells expressing Scd2-GFP from its native promoter treated with 0.1 μg/ml P-factor. Cells were grown in MSL-N medium for 4 hr at 30 ºC before imaging. (Right click to download).
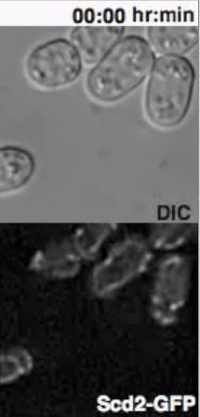
Supplemental Movie 5: Deconvolved single z-plane epifluorescence and DIC timelapse of h-sxa2 cells expressing Scd2-GFP from its native promoter treated with 1 μg/ml P-factor. Cells were grown in MSL-N medium for 4 hr at 30 ºC before imaging. (Right click to download).
| YSM995 | h- wt |
| YSM1371 | h+ wt |
| YSM 1396 | h90 wt |
| YSM2534 | h- myo52-3GFP::kanMX |
| YSM2730 | h+ myo52-tdTomato::natMX Pmap3:GFP::ura4+@ura4locus |
| YSM2731 | h- scd2-GFP::natMX sxa2∆::kanMX |
Table S1: Strains Used in This Study.
