A wild type adult ICR (CD1) mouse heart typically yields 500,000 to 1 million cardiomyocytes from a successful isolation. Immediately after isolation, the cells maintain a mostly rod-shaped appearance (Figure 3A) with intact sarcomeres and can be used for functional studies involving cardiomyocyte contractility. A high percentage of rod-shaped cardiomyocytes (above 90%) is an indication of effective perfusion and digestion. Viable cardiomyocytes will be large (~ 100 – 200 μm in length) and appear to have a sharp outer membrane under brightfield illumination (Figure 3A). Immunostaining for cardiomyocyte-specific sarcomeric markers, such as alpha-actinin, will result in a distinct sarcomeric banding pattern (Figure 3C). In conjunction with morphological analysis and nuclear staining with DAPI, immunostaining can be used to assess the purity of the isolated cardiomyocytes. Fibroblasts will be small and round just after isolation, but will quickly attach and spread thin on cell culture plastic, thus enabling facile distinction from cardiomyocytes.
A low percentage of rod-shaped cardiomyocytes is an indication of an unsuccessful isolation. This can result from a failure to quickly cannulate the aorta after the heart is extracted from the animal. Inefficient perfusion, due to air embolism or incorrect cannulation for example, can also impact the morphology and viability of the cardiomyocytes.
After several days in culture, the cardiomyocytes assume a rounded morphology (Figure 3B). Within 1 – 2 weeks, live cardiomyocytes attach to the substrate and begin spreading (Figure 3D, E). Dead cells can be identified by trypan blue inclusion. A successful isolation and culture will yield approximately 50% live cardiomyocytes after 1 week. Contamination by non-cardiomyocytes, such as fibroblasts will result in a high percentage of these cells later in culture due to their proliferative potential. This can be avoided by increasing the number of gravity sedimentations and/or through pre-plating to increase the purity.28
Transduction with adenovirus can be used to achieve strong gene expression within 48 – 72 hr (Figure 3E). Adenovirus serotype 5 is efficient at transducing adult mouse cardiomyocytes in vitro due to the high expression of the coxackie-adenovirus receptor, but may depend on the type of media used.20

Figure 1. Cardiomyocyte Isolation Equipment and Instrumentation. (A) Surgical instruments used to extract the mouse heart and cannulate the aorta, from top to bottom: hemostats, tissue forceps, curved forceps, Dumont #5 fine forceps, small dissecting scissors, fine iris scissors, fine squeeze scissors. (B) The Langendorff perfusion system used to digest the mouse heart. Perfusion buffers are aspirated through the inlet tube (1) by the perfusion pump (2) and through the pump outlet tube (3) into the inlet port on the heated water jacket (4). The perfusion buffers are warmed by the heated water jacket as they travel through the spiral glass tube, the debubbling chamber, and then out of the cannulation needle attached to the perfusion outlet port (5), which is connected by a stopcock. The conical tube below catches the perfusion buffers, which are re-circulated through the inlet tube. The pressure port (6), compliance port (7) and vent (8) remain closed during perfusion in order to maintain a constant flow rate. The water jacket is heated by a circulating water bath entering through the jacket inlet (9) and outlet (10) ports. A ring stand is used to hold the heated water jacket in a vertical position (red clamps, black base below the purple conical tube rack). Please click here to view a larger version of this figure.
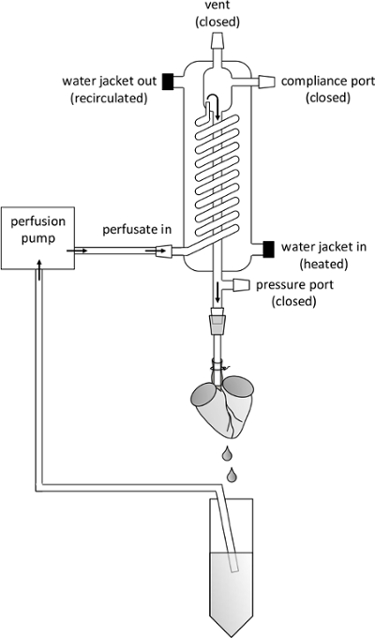
Figure 2. Schematic Overview of Perfusion System. Important Parts of the Perfusion Apparatus are Labeled as Referred to in the Manuscript. The flow direction of perfusion buffer is indicated by arrows. Note the positioning of the aortic cannula, the tip of which should be visible through the translucent aorta, ensuring it is placed above the aortic valve. Please click here to view a larger version of this figure.
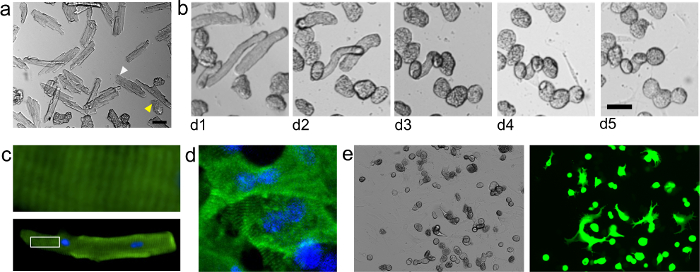
Figure 3. Isolated Adult Mouse Cardiomyocytes. Cardiomyocytes Were Isolated and Seeded onto Laminin-Coated 96 well Plates as Described in the Protocol Herein. (A) Freshly isolated cardiomyocytes, seeded at approximately 8,000 cells/cm2. Note that freshly isolated, viable cardiomyocytes appear approximately rod shaped with sharp edges (white arrowhead). However, cardiomyocytes that appear to have a diffuse outer membrane under brightfield are dead. During the first several days of culture, the cardiomyocytes assume a rounded shape (B). (C) Immunostaining for alpha-actinin (green) reveals characteristic sarcomeric banding in a cardiomyocyte at d1 post-isolation. Nuclear stain (DAPI) shown in blue. Top shows zoomed image of outlined region in lower image. (D) Organized sarcomeres can also be seen after redifferentiation in well spread cardiomyocytes at d11 post-isolation. (E) Brightfield and epifluorescent images show cardiomyocytes at day 11 post-isolation transduced with adenovirus carrying a GFP reporter under the control of a CMV promoter (a gift from Robert Gerard at UT Southwestern). Cells were seeded at approximately 18,000 cells/cm2 and were transduced on day 0 using a multiplicity of infection of approximately 800 plaque-forming units (PFU) per cell. Scale bars: 50 μm. Please click here to view a larger version of this figure.
| Cardiomyocyte isolation buffer (CIB) | |||
| Add 900 ml of ultrapure water to a clean beaker under magnetic stirring. | |||
| Add the buffer components in the table as indicated. | |||
| Component | MW | Final Conc (mM) | 1X (g/L) |
| NaCl | 58.44 | 120 | 7.013 |
| KCl | 74.55 | 5.4 | 0.403 |
| Na2HPO4.7H2O | 268.07 | 0.33 | 0.088 |
| MgSO4.7H2O | 246.48 | 0.5 | 0.123 |
| Taurine | 125.1 | 30 | 3.753 |
| BDM | 101.1 | 10 | 1.011 |
| HEPES | 238.3 | 25 | 5.958 |
| Glucose | 180.16 | 22 | 3.964 |
| Adjust the pH with 5 M NaOH. | |||
| Adjust the volume to 1 L with ultrapure water. | |||
| Sterile filter w/ 0.22 μM vacuum filter. | |||
| Add 0.5 μl of 0.1 U/ml insulin per liter of cardiomyocyte isolation buffer. | |||
| Perfusion buffer | |||
| Component | Final | Volume (ml) | |
| CIB | – | 200 | |
| EGTA (0.4M) | 0.4 mM | 0.2 | |
| Total | – | 200 | |
| Digestion buffer | |||
| Component | Final | Amount | |
| CIB | – | 15 ml | |
| CaCl2 (1M) | 300 μM | 4.5 μl | |
| Protease XIV | 0.2 mg/ml | 3 mg | |
| Collagenase II | 2.4 mg/ml | 36 mg | |
| Total | – | 15 ml | |
| Stop buffer | |||
| Component | Final | Volume (ml) | |
| Perfusion buffer | 95% | 14.25 | |
| CaCl2 (1M) | 1.5 mM | 22.5 | |
| FBS | 5% | 0.75 | |
| Total | – | 15 ml | |
| Culture media | |||
| Component | Final % | Volume (ml) | |
| MEM media | 90% | 90 | |
| FBS | 10% | 10 | |
| Primocin | 0.2% | 0.2 | |
| Total | 100% | 100 ml | |
| Allow culture media to equilibrate at 37 °C, 5% carbon dioxide. | |||
Table 1: Solutions and Buffers.
