Generation of Organ-conditioned Media
An overview diagram/schematic of the process of organ isolation and generation of conditioned media is presented in Figure 1, with representative photographic images of the procedure shown in Figure 2. It should be noted that when this protocol was first under development, liver was included in our analysis because it is a common site of breast cancer metastasis. However, because of the large amount of proteases produced and secreted by the liver, it is very difficult to keep the liver viable long enough to generate good quality conditioned media. Liver-conditioned media is also not stable once isolated, and there tends to be a lot of protein precipitate, likely for the same reason. Therefore liver was not included in any further analysis.
For the generation of lymph node- and bone-conditioned media specifically, the phenotype of isolated stromal cells (LNSCs and BMSCs) can be confirmed by observing their cellular morphology by microscopy combined with immunophenotyping by standard flow cytometry. Stromal cells should be adhesive in culture, with LNSCs demonstrating an elongated and fibroblastic phenotype (Figure 3A) and BMSCs demonstrating a smaller appearance (Figure 3C). Flow cytometric analysis confirms that LNSCs are largely CD45– and gp38+, with ~ 60% of cells possessing a CD45–gp38+ phenotype indicative of LNSCs16 (Figure 3B). The BMSCs should be positive for CD44 and CD29, weakly positive for CD105 and Sca-1, and negative for CD73 (Figure 3D – H), indicative of BMSCs17,18.
Breast Cancer Cell Response to Organ-appropriate Migration Patterns and Gene Expression Changes
Using this ex vivo model system, it has previously been demonstrated that human breast cancer cells with varying genetic backgrounds exhibit differential migration and growth patterns towards specific organ conditions. Notably, these patterns reflect the known metastatic dissemination patterns of these cell lines in vivo13. In the current study, bone (231-BoM)- and lung (231-LM2)-seeking variants of the MDA-MB-231 human breast cancer cell variants (a kind gift from Dr. Joan Massagué11,12) have also been used in a transwell migration assay to confirm organ-appropriate migration patterns. The results demonstrate that the bone-seeking 231-BoM variant preferentially migrates to the bone marrow-conditioned media over brain- and lung- conditioned media (Figure 4A, left). Similarly, the lung-seeking 231-LM2 variant preferentially migrates to lung-conditioned media over bone marrow-conditioned media and brain-conditioned media (Figure 4A, right). Furthermore, RT-qPCR analysis demonstrates that exposure of parental MDA-MB-231 breast cancer cells to bone- or lung-conditioned media causes an upregulation of genes associated with bone-specific metastasis (CXCR4, IL-11, TGFß1) or lung-specific metastasis (VCAM1) of breast cancer cells in vivo11,12 (Figure 4B). These results demonstrate the validity of the ex vivo system with regards to the expected organ tropism of breast cancer cells in vivo, and indicates that soluble factors present in organ-conditioned media can influence metastatic cell phenotype in addition to promoting migration.
Organ-conditioned Media Contains Potential Soluble Mediators of Metastatic Behavior
A biotin label-based mouse antibody array was used to assess the presence and identity of common soluble factors in organ-conditioned media from different organs. This array includes antibodies against 308 of the most common soluble mouse proteins.This protein array has previously been used to identify soluble factors of interest associated with the metastatic cascade present within the lung and bone marrow conditioned media as shown in Chu et al (2014) and Pio et al (2015) respectively13,14. In the current study, protein array results are shown for basal media (Figure 5A) compared to lymph node- (Figure 5B) and brain-conditioned media (Figure 5C). Highlighted beside each array are soluble factors present in the media that have been found to be associated with steps in the metastatic cascade in the literature19-29. These soluble factors may be worth investigating as potential mediators of secondary metastases within these organ sites.
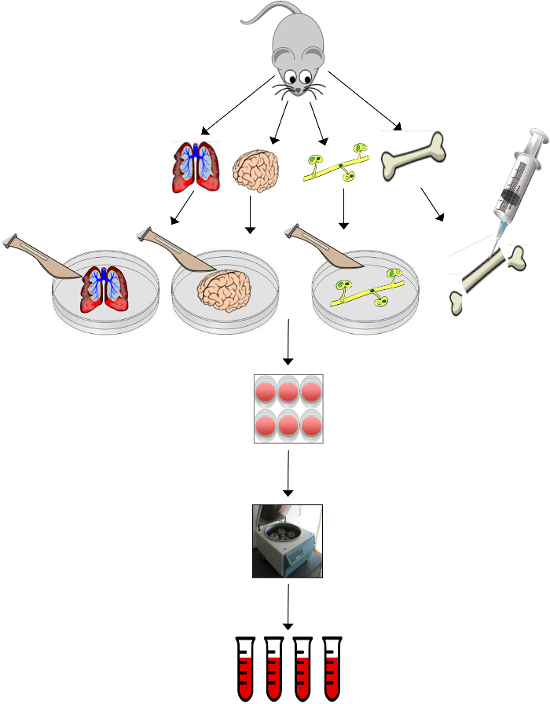
Figure 1. Overview Schematic of the Process of Organ Isolation and Generation of Conditioned Media. Tissues (lungs, brain, femur, tibia and lymph nodes) are harvested from the mouse. Lungs, brain and lymph nodes are minced with a scalpel. Bone marrow is harvested from the medullary cavity of the bones by pushing PBS through the cavity with a 27½ G needle. Lung and brain tissue fragments are incubated for a period of 24 hr before diluting with three equivalent volumes of fresh basal media and collected to make lung- and brain-conditioned media. Bone marrow and lymph node stromal cells are passaged 2 – 3 times before incubating with basal media to make bone and lymph node-conditioned media. Harvested media can be pooled and stored at -80 °C until ready for use in downstream assays such as protein arrays, cellular growth assays, cellular migration assays, tumorsphere formation, and RT-PCR. Please click here to view a larger version of this figure.

Figure 2. Representative Photographic Images of the Procedure for Organ Isolation and Generation of Conditioned Media. At Day 0, the lungs, brain, femur, tibia and lymph nodes are collected from the mouse. The lungs, brain and lymph nodes are placed in a 60 mm2 glass petri dish and minced with two scalpel blades to approximately 1 mm3 in size. The bone marrow is harvested from the medullary cavity of the bones by pushing PBS through the cavity with a 27 ½ G needle into a fresh tube of PBS. The lung and brain tissue fragments are incubated for a period of 24 hr at 37 °C and 5% CO2 before diluting with three equivalent volumes of fresh media and collected to make lung- and brain-conditioned media after 24 hr. The bone marrow and lymph node stromal cells are incubated at 37 °C and 5% CO2 and passaged 2 – 3 times (~ 3 weeks total) before replacing media with serum-free basal media to make bone and lymph node-conditioned media. Please click here to view a larger version of this figure.
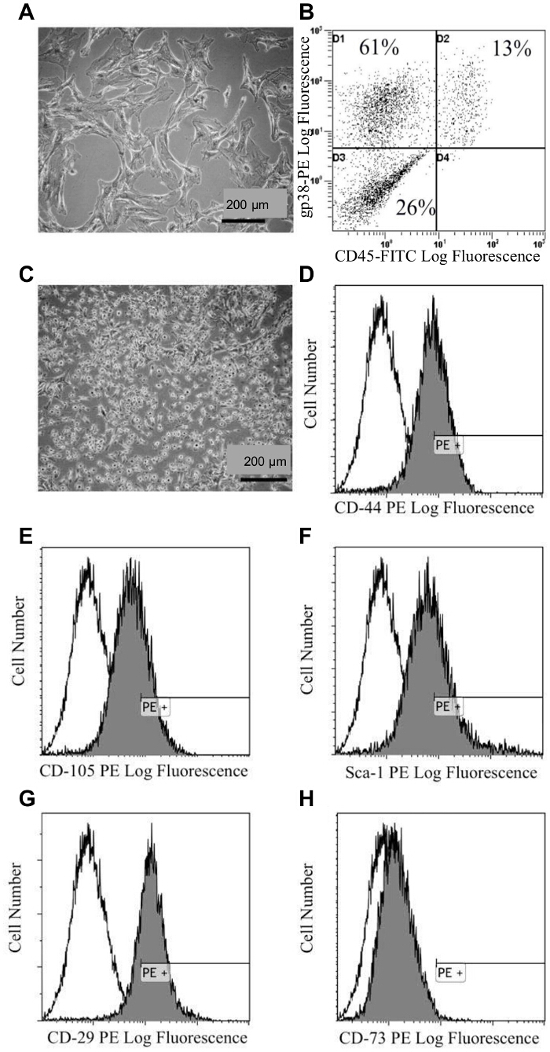
Figure 3. Characterization of Isolated Lymph Node and Bone Marrow Stromal Cells. (A) Bright-field microscopy image showing the morphology of lymph node stromal cells (LNSCs). (B) Representative flow cytometry analysis of LNSCs using a phycoerythrin (PE)-conjugated gp38 antibody and a fluorescein isothiocyanate (FITC)-conjugated CD45 antibody. (C) Bright field microscopy image showing the morphology of bone marrow stromal cells (BMSCs). (D-H) Representative flow cytometry analysis of BMSCs using PE conjugated (black profiles) antibodies against (D) CD44, (E) CD106, (F) Sca-1, (G) CD29, (H) CD73 antibodies relative to the isotype control (white profiles). A minimum of 10,000 viable events were collected per sample. This figure has been modified from 13. Please click here to view a larger version of this figure.
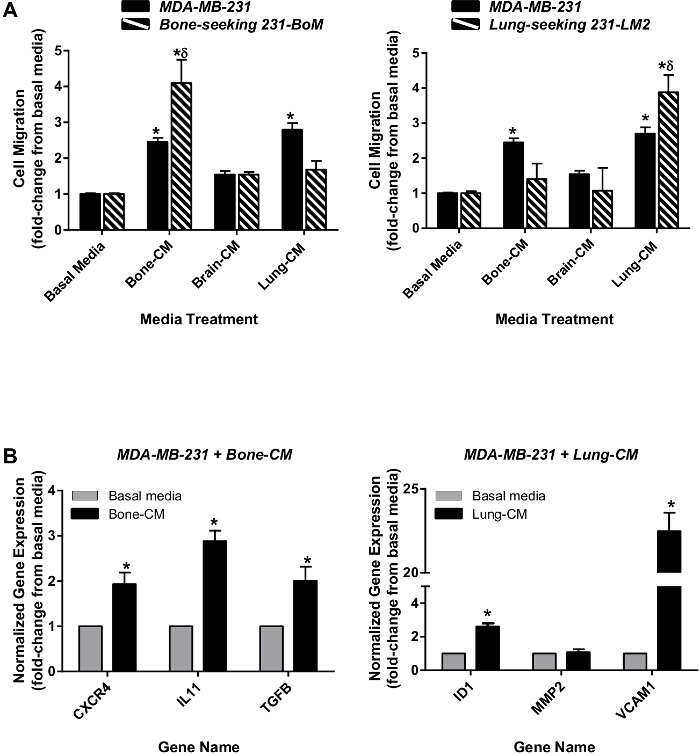
Figure 4. Organ-appropriate Migration Patterns and Gene Expression Changes in Response to Organ-conditioned Media. (A) Bone-seeking 231-BoM variants (left, striped bars) or lung-seeking 231-LM2 variants (right, striped bars) of the MDA-MB-231 human breast cancer cell line as well as parental MDA-MB-231 cells (both panels, solid bars) were plated onto gelatin-coated inserts (with 8 µm pores) prior to placement in basal media (DMEM:F12+ concentrated mitogen supplement) or organ-CM. Plates were incubated at 37 °C and 5% CO2 for 18 hr. (B) Whole population parental MDA-MB-231 cells were exposed to either bone-CM (left) or lung-CM (right) for 48h. RNA was isolated and quantified using RT-qPCR. Gene expression changes in response to organ-CM (black bars) were normalized to GAPDH and expressed relative to baseline expression in basal media (grey bars). Data are presented as mean ± SEM (N = 3; fold-change from respective negative control of basal media). * = significantly different from basal media; δ = significantly different from parental cell line treated with the same media conditions; P <0.05, ANOVA. Please click here to view a larger version of this figure.
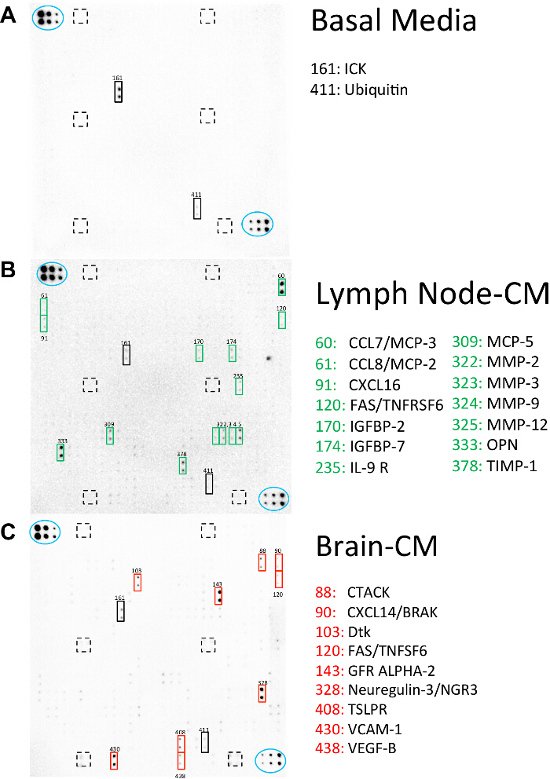
Figure 5. Organ-conditioned Media Contains Potential Soluble Mediators of Metastatic Behavior. A biotin label-based mouse antibody array was used to identify soluble factors present in lymph node-CM and brain-CM. Membranes were incubated with biotinylated samples of (A) basal media (DMEM:F12+ concentrated mitogen supplement), (B) lymph node-CM or (C) brain-CM at 4 oC O/N and visualized using chemiluminescence. Blue ovals indicate internal positive controls, lined boxes indicate internal negative controls and solid boxes indicate soluble factors of interest that are potentially involved in the metastatic behavior of breast cancer cells to the lymph node or brain. Please click here to view a larger version of this figure.
