Decellularization
On average, the time to decellularization of a P3 heart using this protocol is approximately 14 hr. given an average heart weight of 23 mg for the P3 neonate.
Acellularity
Figure 3a demonstrates a fully intact P3 neonatal heart (whole mount). Figure 3b shows the same heart following decellularization. Figures 4a and 4b show the hematoxylin and eosin staining of intact and decellularized hearts, respectively. Note the absence of hematoxylin positive nuclei and the diminution of eosinophilic structures in the decellularized heart. In addition, the DNA content of the decellularized heart is significantly reduced from 68.08 ± 2.25 µg in the intact heart (n=6) to 4.73 ± 2.27 µg in the decellularized heart (n=5).
Collagen Immunoreactivity
The maintenance of the extracellular matrix (ECM) following decellularization is essential to repopulation with exogenous cells and to the functionality of the matrix. To evaluate the content of the neonatal ECM in intact and decellularized ECM, immuno-staining for collagen IV was performed. Figure 4c and d demonstrate that collagen IV is robustly expressed in both the intact and decellularized heart and that the localization of this protein is maintained following cell removal, while DAPI positive nuclei are effectively removed (Figure 4e-h).
DNA Content
The presence of DNA is used as an additional indication of cellularity. In Figure 5, DNA is demonstrated to be decreased by 93% in neonatal hearts following detergent based decellularization. This degree of DNA reduction is consistent with reports in the literature using detergent based decellularization in other tissues15-16.
Recellularization
We have performed immunohistochemistry to determine the expression of various cardiomyocyte markers in the recellularized heart. Figure 6 illustrates cells that have migrated into the wall of the left ventricle (Figure 6A and 6B). Figure 6C demonstrates the DAPI labeling of the recellularized heart. Figure 6D-G illustrates cells that are positive for NKX 2.5, mCherry, α-actinin and DAPI, respectively. NKX2.5 is known to label cardiac progenitor cells, whereas α-actinin is a sarcomeric protein that marks differentiated cardiomyocytes. We have observed a majority of cells which express all of these markers (pink; Figure 6H), indicating that these cells continue to express cardiomyocyte markers even after 23 days of perfusion.

Figure 1. Schematic of decellularization hardware. A. 60 cc syringe barrel reservoir for detergent solution. B. Catheter assembly with irrigation syringe as detailed. C. Detail of the drawn PE tubing catheter tip. D. Decellularization chamber and septum with drain. Please click here to view a larger version of this figure.

Figure 2. Schematic representation of the heart bioreactor. 1. Humidification of carbogen gas (Green lines represent gas flow). 2. Peristaltic pump drive for media perfusion and oxygenation (Purple lines represent media flow through oxygenator). 3. Thin wall oxygenator. 4. Sheet oxygenator and media reservoir. 5. Preload chamber and bubble trap. 6. Heart chamber (Red lines represent media flow to and from the heart). Please click here to view a larger version of this figure.
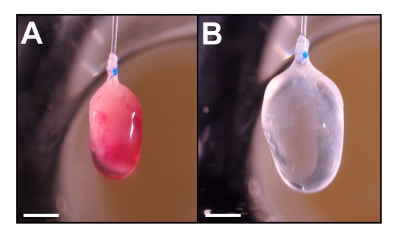
Figure 3. Whole mount of P3 neonatal mouse heart before (A) and after decellularization (B). This heart is decellularized using the Langendorff perfusion method as described in Method 2. Note that the heart becomes translucent and slightly enlarged following perfusion (B). Scale = 2mm. Please click here to view a larger version of this figure.

Figure 4. Histology of native and decellularized P3 neonatal mouse heart. Cryostat section (10 µm) were stained with H&E (A, B), Collagen IV (C, D, G, H) and DAPI (E, F, G, H). Merged images are represented in G and H. H&E staining shows an absence of cell nuclei and cytoplasm in decellularized tissue (B) when compared to the native heart (A). While the Collagen IV content remains following decellularization (D), DAPI staining (a marker of nuclei) is abolished. The merged images demonstrate the colocalization of Collagen IV and DAPI in the naïve heart (G) and the absence of this colocalization in the decellularized heart (H). These data indicate that cells no longer populate the collagen matrix of the heart. Scale = 500 µm. Please click here to view a larger version of this figure.

Figure 5. Evaluation of DNA content. Control (n=6) and decellularized (n=5) hearts assayed for DNA content by the pico-green method. Quantitation expressed as µg of DNA per heart ± standard deviation. Asterisk indicates p<0.01 compared to control. These data indicate that decellularization reduces DNA content significantly in the P3 neonatal heart. Please click here to view a larger version of this figure.

Figure 6. Histology of P3 heart matrix 23 days following recellularization with P1 mCherry expressing cardiomyocytes. Stained with H&E (A, scale = 250 µm, B, scale = 50 µm), DAPI (C, scale = 250 µm), NKX 2.5, mCherry, α-actinin, DAPI, and merged (D, E, F, G, H, scale = 50 µm). We have demonstrated the effective repopulation of the collagen matrix with P1 cardiomyocytes (A-C). Additionally, we observed that m-cherry positive cardiomyocytes express Nkx2.5 and α-actinin 23 days following introduction into the collagen matrix. These data indicate that these cells maintain their cardiomyocyte identity for extended periods of time. Please click here to view a larger version of this figure.
